





Protected areas can prevent forest loss, but their effects on forest fragmentation and forest regrowth are poorly understood. Furthermore, the importance of protected areas in shaping these forest spatial changes may depend on different socioeconomic drivers (e.g. population size, distance to cities, proportion of local people working in non-farm occupation), but the empirical evidence on such dependence is very scarce. Here, we used contra factual technics to assess whether biosphere reserves (n = 19) in the Mesoamerican biodiversity hotspot can reduce forest loss and fragmentation and promote forest regrowth during the period 2000–2020. We used satellite imagery and governmental data to assess the socioeconomic factors driving these changes. Particularly, using multimodel inference analysis, we tested whether higher non-farm occupation, combined with low demographic pressures, reduces forest loss and fragmentation and promotes forest regrowth. We found that reserves reduce forest loss and preserve less-fragmented configurations, however, they neither reduce fragmentation rate nor promote forest regrowth. Forest loss rate inside the reserves decreased as non-farm occupation enhanced and the density of rural settlements decreased. Therefore, promoting higher opportunities in non-farm economic activities and planning rural settlements distribution around reserves could help to increase the effectiveness of reserves for forest conservation.
Forest loss is considered a major driver of biodiversity loss (Watling et al., 2020). This process is particularly acute in tropical forests (Hansen et al., 2013). Forest loss is also causing the fragmentation of tropical forests worldwide (Taubert et al., 2018), which can increase the susceptibility of remaining patches to post-fragmentation threats such as negative edge effects, logging, hunting, and fires (Malhi et al., 2014). To prevent this global pattern of tropical forest degradation, protected areas (PAs) are frequently considered a major strategy for biodiversity conservation since they can reduce deforestation (Spracklen et al., 2015) and protect species.
Despite their conservation value, PAs face a growing threat due to human activities (Jones et al., 2018), which result in forest cover changes inside and outside them (Wade et al., 2020) impacting their conservation. Thus, forest cover changes may affect PAs conservation in several ways. On the one hand, forest loss is identified as the main cause of biodiversity loss (Newbold et al., 2015). On the other hand, despite that forest fragmentation may have negative effects (Ewers and Didham, 2006), most of them are neutral or positive (Fahrig, 2017), nonetheless this is currently under debate (Fletcher et al., 2018). Furthermore, forest regrowth in the tropical forest promotes the recovery of biodiversity (Rozendaal et al., 2019). Thus, evaluating the pattern of forest cover changes in PAs is critical to improving management and conservation strategies.
Understanding the causes of forest cover changes in PAs is also of paramount importance. Overall, two types of drivers can be distinguished: proximate and underlying. The proximate drivers correspond to human activities that directly modify the forest cover, such as infrastructure extension, agricultural expansion (both subsistence and large scale), and wood extraction (Curtis et al., 2018). Environmental factors, such as hurricanes, fires, landslides, severe droughts can also act as proximate drivers of forest cover changes (Geist and Lambin, 2001). The underlying drivers are related to factors that indirectly modify the forest cover by modulating proximate drivers, such as demographic, economic, technological, political-institutional, and cultural factors (Geist and Lambin, 2001). For example, demographic-related factors, such as human population growth and population density, are recognized as major drivers of deforestation (Aide et al., 2013) as they are linked to forest resource consumption. Economic factors also play a critical role in forest cover dynamics. Access to markets is a key factor influencing trade prices for agricultural products, which in turn affects deforestation (Angelsen, 2010).
Protected areas in the tropics are usually inhabited by rural communities that subsist from agriculture activities. These communities derive low income from agriculture due to the high costs of product transportation (Angelsen, 2010), which may increase deforestation to expand the cultivated area seeking higher income (Ferraro et al., 2011). Also, in tropical regions, a shift from economies based on agriculture to non-agricultural economies has been indicated as one of the drivers producing a decrease in forest loss rates and an increase in forest regrowth, a process known as forest transition (Meyfroidt and Lambin, 2011). According to the forest transition theory, non-agricultural economies reduce the pressure on forests because local people obtain their income from activities representing less change in land use (Rudel et al., 2005). For example, rural communities having labor opportunities on tourism (Hoang et al., 2014) and industrial activities (Wunder, 2003) carry out less deforestation than communities depending only on agriculture (Kovacic and Viteri Salazar, 2017). However, the effect of deforestation reduction, associated with the emergence of non-farm occupation in tropical regions, on PAs conservation function is still an open issue, and empirical evidence is very scarce.
Here, we first apply contrafactual matching methods to test the hypothesis that PAs not only prevent forest loss but also fragmentation and promote forest regrowth during the period 2000–2020. Second, we determined the effect of some underlying socioeconomic drivers on forest cover changes within some reserves located in Mesoamerica. We hypothesize that economies based on non-farm occupation, combined with low demographic pressures, reduce forest loss and fragmentation, and promote forest regrowth inside reserves.
MethodsStudy systemWe selected all Mexican MAB-UNESCO Biosphere reserves (hereafter reserves) located in the Mesoamerican hotspot, which were established before the year 2000 (Fig. 1, Table S1, Appendix 1). We exclude from our analysis the biosphere reserves La Encrucijada and Pantanos de Centla because of the low performance of forest/no forest identification (see below), therefore we include 19 reserves. The reserves show a gradient of forest loss, with the remaining forest cover within the reserves ranging from 36% to 98%. We combined the Lacan-Tun/Montes Azules and Los Petenes/Ria Celestún reserves into two reserve complexes called Lacandona and LPRC, respectively, because they shared borders, same biome, and management.
Land cover classification
To characterize forest changes inside reserves and unprotected areas (areas in Mexican tropic non included in any PA, see Fig. 1), we performed a supervised classification of Landsat images for the years 2000 and 2020. We used three major classes: forest, no-forest, and water. We evaluated the accuracy of this classification using a set of independent class cover validation points and determined that the mean (±SD) value of the overall satellite image classification accuracy was 94.4% ± 0.02 for the 2000 year and 94.4% ± 0.04 in 2020 (Table S4, Appendix 2). We conducted a cover change analysis to identify the areas with forest loss, forest regrowth, and no change for a period of 20 years, from 2000 to 2020 in the whole studied area.
We also calculated forest loss rate, forest regrowth rate, and forest fragmentation rate within reserves. To calculate forest loss rate (FLR) and forest regrowth rate (FRR) we use the next formula:
where A0 is the area covered by forest at 2000 (t0) and t1 represents the year 2020. FLA is the area covered by forest at t0 but no forest at time t1 while FRA represents the area with the opposite transition. We quantified the number of forest patches in the year 2000 (NP2000) and the year 2020 (NP2020) and calculated forest fragmentation rate (FFR) with the formula:Matching analysis for forest loss, forest regrowth, and fragmentation data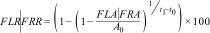
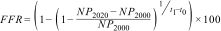
Protected areas are often located in isolated places with steeper slopes, higher elevations, and poorer soils than unprotected places (Joppa and Pfaff, 2009). Thus, a simple comparison of forest changes between protected and unprotected regions may overestimate the effect of legal protection (dos Santos Ribas et al., 2020). Matching analysis is a quasi-experimental approach that statistically allows comparing sampling units with similar covariates and, therefore, to test the effect of treatment without the effect of covariates (Ho et al., 2011). This technic matches sampling units of treatment and control groups with similar covariates values to test the effect of a treatment. Matching technics have been used previously to assess the effect of PAs on forest loss (Yang et al., 2021), but its use regarding forest regrowth and forest fragmentation in PAs is less frequent (but see Sims, 2014).
We used two different sampling strategies for the matching analysis: the first with one million points uniformly seeded in the entire studied area (both inside and outside reserves) at 1 km of separation, and the second with 1500 circular microlandscapes with 3 km of radius randomly seeded in the same area (see Fig. S1 and S2, Appendix 1). Each sampling point has information regarding whether they are located inside reserves or in unprotected areas, and about the changes in forest cover that occurred during 2000–2020 in its specific location (i.e. forest loss, forest gain, or no change). We used the microlandscapes to document forest fragmentation inside reserves and in unprotected areas using formula 2. Furthermore, each sampling unit had the value of the three covariates that may act as confusing factors on forest cover changes: distance to cities, distance to roads, and agriculture suitability (see Appendix 2). To properly compare forest cover changes in reserves and unprotected areas we only include those samples located within forest cover in the year 2000 for the forest loss data (n = 423,900), and within no forest cover in 2000 for the forest regrowth data (n = 331,623).
We estimated propensity scores for forest loss, forest regrowth, and fragmentation data using binomial error distribution with a logit function where the response variable was the treatment (protected or unprotected). After matching, all standardized mean differences for the covariates were below 0.1 which indicates a good balance between control and treatment samples (see Appendix 3). The matching analysis was performed using the MatchIt package (Ho et al., 2011) of R (R Core Team, 2021).
Underlying drivers of forest cover changeAs underlying drivers of these forest changes, we considered two economic indicators: (i) the distance to major cities (localities with a population larger than 15,000 people) as a proxy of access to markets, and (ii) the non-farm occupation (i.e. the mean proportion of people, of the municipalities surrounding the biosphere reserves, working in the professional, services, and industrial sectors; Table S2, Appendix 1) in the year 2000. Population growth rate (during 2000–2020), population density in the year 2000, and rural settlement density in the year 2000 were also used as indicators of demographic factors. Using a correlation and a collinearity analysis, we proved that these indicators were not correlated (see Fig. S3, Appendix 1). During the first stage of our study, we considered including other indicators, such as unemployment rate, marginalization, human development, and agricultural subsidies. However, they were not included in our analysis because of the high correlation with the selected indicators (r > 0.5, p < 0.05, Variance Inflation Factor (VIF) > 2). Most selected indicators of underlying drivers are common in other studies (Aide et al., 2013; Figueroa et al., 2009). The theoretical relationship with forest loss and details of the calculation of these indicators can be found in Table S3 of Appendix 1 and Appendix 2, respectively.
Data analysisWe tested for the effect of reserves (i.e., protection) on forest loss, forest regrowth, and forest fragmentation. For the first two variables, we used generalized mixed models with binomial error distribution for each one. We included forest change as binary response variables (forest loss/no change and forest regrowth/no change) and the protection condition (inside reserves or unprotected) as a fixed effect factor. To test for the effect of the protection condition on forest fragmentation rate and the number of patches in the year 2020, we used a generalized linear mixed model with a Gaussian error and a Poisson error, respectively. We included the biome type as a random effect. We performed the models with pre-matching and matching data. Matched data models were weighted using the weights automatically calculated by the matching algorithm that accounts for the potential differences in the number of samples in each treatment.
To determine the effect of the indicators of socioeconomic drivers on the forest loss, fragmentation, and forest regrowth rates, we used a multimodel inference approach with linear models (Burnham and Anderson, 2002). Because of the small sample size (n = 17), we limited to 3 the maximum number of terms in the models to avoid overfitting. Therefore, for each response variable, we constructed all models that represent all possible model combinations with 1, 2, and 3 predictors and their pair interaction (i.e., approximately 600 models). To calculate standardized parameter estimates (i.e., slopes), we applied a z-score transformation for all predictor variables. We calculated the Akaike Information Criterion corrected for small samples (AICc), and the model with the lowest AICc value was considered the best model. The percentage of deviance explained by the best model was calculated with square-R (goodness-of-fit). The effect size of each variable was estimated with the model-averaged standardized parameter estimates.
ResultsThe study biosphere reserves had significantly less probability of forest loss than unprotected places, even when controlling for any bias caused by differences in environmental conditions. Our models indicated that reserves reduced forest loss by about 54% compared to unprotected places (Fig. 2a). Regarding fragmentation, we found that reserves have a significantly smaller number of patches in 2020 than unprotected places (Fig. 2b). Nonetheless, reserves did not reduce the fragmentation rate (Fig. 2c). Reserves had a higher forest regrowth probability than unprotected places without considering the differences in environmental conditions. However, reserves had a similar forest regrowth probability than unprotected places when we controlled for covariates (Fig. 2d).
 estimated forest loss probability (the probability that a pixel transit from forest cover to no forest cover in the period 2000–2020, (a), the estimated number of patches in the year 2020 (b), estimated fragmentation rate (annual relative change in the number of patches) in the period 2000–2020 (c), and estimated forest regrowth probability (the probability that a pixel transit from no forest cover to forest cover in the period 2000–2020, (d) in the reserves and unprotected zones before and after the matching analysis. ns: non-significant, *p < 0.05, **p < 0.01, ***p < 0.001.")
Mean (±SE) estimated forest loss probability (the probability that a pixel transit from forest cover to no forest cover in the period 2000–2020, (a), the estimated number of patches in the year 2020 (b), estimated fragmentation rate (annual relative change in the number of patches) in the period 2000–2020 (c), and estimated forest regrowth probability (the probability that a pixel transit from no forest cover to forest cover in the period 2000–2020, (d) in the reserves and unprotected zones before and after the matching analysis. ns: non-significant, *p < 0.05, **p < 0.01, ***p < 0.001.
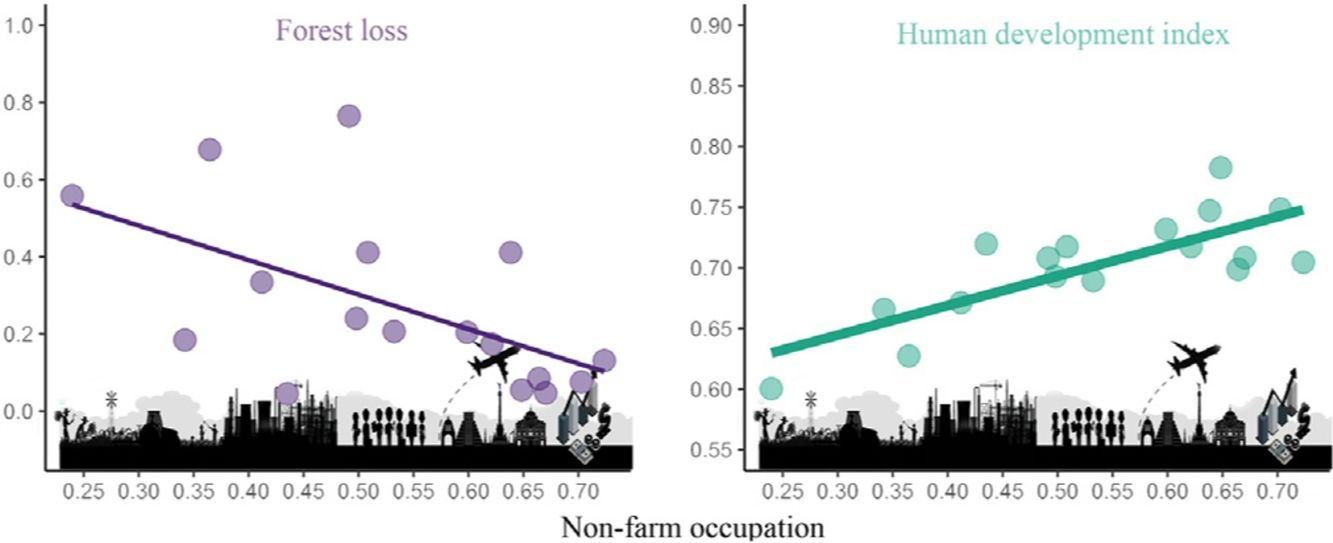
The forest loss rate inside the reserves was strongly and negatively related to non-farm occupation and positively to the density of rural settlements (Fig. 3a). In addition, the fragmentation rate in the reserves was strongly and positively associated with the population growth rate (Fig. 3b). Finally, the forest regrowth rate was mostly and positively influenced by the density of rural settlements and negatively by the population growth rate (Fig. 3c).
. For practical reasons we only show the six best models according to the AICc. The vertical axis shows the underlying driver indicators, and the horizontal axis shows the model-averaged standardized parameter estimates, a proxy of effect size. Bars oriented to the right represent positive responses, while bars oriented to the left are negative responses. In all the cases the explanatory variable has a higher model-averaged standardized parameter estimates value than mean unconditional variance, and therefore statistical confidence. The percentages in each panel indicate the goodness-of-fit of the best model.")
Response of forest spatial changes assessed inside the biosphere reserves to different underlying drivers (showed in different fill colors). For practical reasons we only show the six best models according to the AICc. The vertical axis shows the underlying driver indicators, and the horizontal axis shows the model-averaged standardized parameter estimates, a proxy of effect size. Bars oriented to the right represent positive responses, while bars oriented to the left are negative responses. In all the cases the explanatory variable has a higher model-averaged standardized parameter estimates value than mean unconditional variance, and therefore statistical confidence. The percentages in each panel indicate the goodness-of-fit of the best model.
This study assesses the patterns and underlying drivers of land-use change in 19 reserves within the Mesoamerican biodiversity hotspot. Our findings confirm the key role played by protected areas in preventing forest loss within reserves. However, we go further than previous studies (Joppa and Pfaff, 2011; Spracklen et al., 2015; Yang et al., 2021) by showing that reserves had less fragmented landscapes, although the rate of fragmentation was not different from unprotected places. Overall, our results indicate that a high non-farm occupation, and a low demographic pressure, increase the effectiveness of reserves in preventing forest loss. As argued below, these findings have important applied implications for promoting the effectiveness of protected areas in the Mesoamerican biodiversity hotspot.
Do reserves prevent forest loss and fragmentation and promote forest regrowth?Our findings confirm the importance of reserves as key tools to prevent forest loss and to preserve landscapes with a low fragmentation degree. Nevertheless, we did not find evidence supporting the idea that reserves also reduce the forest fragmentation rate. In other words, biodiversity within reserves cope with a lower fragmentation degree than in unprotected zones, but fragmentation increases at similar rates within reserves and unprotected zones. Since forest loss has higher deleterious effects on biodiversity than fragmentation (Fahrig, 2017), our findings represent less concern news for biodiversity inside reserves, without neglecting the potential negative effects of fragmentation (Ewers and Didham, 2006).
Surprisingly, we did not find support to the hypothesis that reserves promote forest regrowth. This contrasts with other works that suggest a positive effect of protected areas on forest recovery (Borda-Niño et al., 2020). This apparent contradiction may be clarified by our matching analysis. In our study, a simple comparison of protected and unprotected areas, without controlling for environmental covariables (pre-matching condition), supports the expected higher forest regrowth within reserves. However, controlling for environmental covariables obliterated such differences. Therefore, the forest regrowth inside and outside reserves does not seem to depend on the establishment of the reserves per se, but on the distance to cities and agriculture suitability (Fig. S4, Appendix 1). Thus, forest regrowth occurs in areas where agricultural activities are not favored because of the biophysical environment (i.e. steeper slopes, high elevations, and poor soils) or because of economic causes (a large distance to markets promote land abandoning and forest regrowth).
Underlying drivers of forest cover changesOur findings indicate that the forest loss rate inside the reserves is mainly related to two main underlying drivers, positively to the density of rural settlements and negatively non-farm occupation. These results suggest that the lack of job opportunities in rural areas, beyond those offered by the agricultural sector, accentuates deforestation as the demand for agricultural land increases in the absence of other livelihood options. As reported by previous studies, reduction of forest loss rates is associated with the increase of non-farm labor activities (Hoang et al., 2014; Wunder, 2003), since these allow local people to obtain revenues without the need of causing more deforestation for arable land (Curtis et al., 2018; Kovacic and Viteri Salazar, 2017; Vedeld et al., 2007). In parallel with these studies, we found that reserves embedded in municipalities with greater labor opportunities in non-farm sectors, especially in the industrial activities (e.g., manufactures, electricity, or construction, see Table S4, Appendix 1), endure lower forest loss rates (Goers and Lawson, 2009). Also, a higher human development index was associated with a greater non-farm occupation (Fig. 4). This finding concurs with studies pointing out that non-farm activities allow local people to obtain higher profits and deal with poverty (Haggblade et al., 2007). According to our study, the emerging non-farm occupation may promote a “win-win” scenario with a positive balance between conservation and development, especially considering that an incipient forest transition process in southern Mexico has been identified (Vaca et al., 2012). However, it is important to note that this “win-win” scenario is likely limited to regions dominated by subsistence agriculture, such as the study reserves in Mexico (Bonilla-Moheno and Aide, 2020; Curtis et al., 2018). Additional studies are required in places where large-scale agriculture oriented to the production of commodities occurs, as the diversification of labor activities could have a relatively weak effect on deforestation rates in contexts where agriculture is typically supported by large capitals.
, and human development index (HDI, (b). Each point represents a single reserve, lines represent the generalized linear model and the grey shadow is the 95% confidence interval.")
As expected, the population growth rate was a strong predictor of forest cover changes, being positively related to forest fragmentation rate, and negatively associated with forest regrowth. Human population size is considered a major driver of forest loss (Aide et al., 2013) and fragmentation (Li et al., 2010). Human pressures such as logging and agriculture are expected to increase with increasing the population size (Ehrlich and Holdren, 1971), which in turn can promote the loss and fragmentation of the remaining forest, and limit the recovery of the forest in degraded lands. Our findings contrast with those of Borda-Niño et al. (2020), which suggest that the forest regrowth rate can increase with increasing (not decreasing) demographic factors such as the density of rural settlements. This counterintuitive finding can be related to the environmental policies after the establishment of the reserves. In Mexico, for example, the creation of reserves often requires the relocation of rural communities that were within the limits established for the reserve, which leads to farmers abandoning their cropland allowing the forest to regrow (Fig. S5, Appendix 1). We infer that a higher forest regrowth occurred especially in reserves that had a higher density of rural settlements in their surroundings. Indeed, previously to the year 2000, when some reserves have not been legally established, the amount of forest loss rate inside reserves was significantly and positively related to the density of rural settlements (R2 = 0.4, P < 0.01, Fig. S6, Appendix 1). Although difficult to support with empiric data, another possibility is that there exist active restoration efforts, as occur in some regions (Juan-Baeza et al., 2015), potentially associated positively with demographic factors (the larger the population, the greater the possibility of restoration).
Finally, we have some caveats about our work to enunciate. Our work assessed the influence of socioeconomic variables measured at the municipality on forest spatial changes, however, there exists the possibility that a finer scale (e.g. locality) outcome more reliable results, nonetheless, this is still a challenge since some variables are not available at fine resolution. In addition, there exist other underlying and proximate drivers (e.g. environmental factors) non-assessed in our work that influence forest spatial changes in protected areas that may need specific hypothesis tests in future studies. On the other hand, since here we assessed forest spatial changes in 20 years, our work may not identify properly early stages of forest regrowth (mostly given by small trees and shrubs) and therefore, underestimates forest regrowth and forest fragmentation.
Concluding remarksOur results indicate that promoting some underlying drivers, specifically increasing labor opportunities in non-farm sectors (i.e. reducing local people’s dependence on agricultural activities) and reducing demographic pressures, could strengthen the conservation function of biosphere reserves by reducing forest loss inside reserves. However, reducing the number of people in an area is still a major challenge in terms of public policies. Therefore, allocating resources to provide more economic opportunities seems to be a better option. In our view, in the absence of further job opportunities, farmers are forced to continuously expand the agricultural frontier. Finally, it is important to note that increasing non-farm jobs opportunities should be considered in the management plans of reserves, as a way to mitigate negative anthropogenic effects on them.
Conflict of interestThe authors have no conflicts of interest to declare.
Data and code availabilityData of spatial changes inside the reserves, methodological workflow, and matching algorithm code are available online https://github.com/aulizdaniel/Biosphere-resereves-data.
FundingThis research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
This research is part of the doctoral project of DAO. DAO thanks Posgrado en Ciencias Biológicas of UNAM and CONACyT for the doctoral scholarship awarded. DAO thanks to Dr. Victor Sánchez Cordero for help and support in the first steps of this research. DAO also thanks Francisco Mora for the technical advice in the development of the research, German Wies for general comments, Dallas Levey for the revision of a previous version manuscript, and Jonathan Auliz for the help in the design of the graphical abstract and conceptual framework. We thank the three anonymous reviewers for their valuable comments which contributed to significantly improving our work.
The following are Supplementary data to this article:





