




Logging is one of the main causes of biodiversity loss in tropical forests. In the past decades there was an increase in the number of studies on the effects of logging on biodiversity, but there has been little advancement for bats, despite their ecological importance. We present a review of studies on the effects of logging on bats in tropical forests worldwide carried out in the past three decades. We aimed at answering the following questions: What is known about the effects of logging on the bat fauna of tropical forests? What are the gaps of knowledge that can be filled? We conducted a literature search of studies on the effect of logging on the bat fauna in tropical forests in the past decades. We surveyed the databases Web of Science, Google Scholar, and Scopus with different keyword combinations: “Bats OR Chiroptera”, “Logging”, “Selective logging”, “Timber extraction”, “Tropic”, “Forest”, and “Tropical Forest”. We found 22 studies focused on Latin America and Southeast Asia. Most studies (81.8%) only compared bat richness and abundance between logged and unlogged areas, where frugivorous bats responded positively to logging, whereas gleaning animalivores bats responded negatively. Few studies (18%) tried to understand how environmental variables, such as changes in vegetation structure, affect bat diversity. We emphasize that future studies aimed at checking the effects of logging on bats should use more than one sampling method in order to obtain more representative samples. Planning should be done with more caution, in order to avoid pseudoreplication and obtain more solid results. Poorly studied regions that have intensive logging, such as the Amazon and the tropical forests of Africa, should receive more attention.
Until 2050 the global demand for timber should triplicate due to the human population growth and the increase in the use of bioenergy, which uses the biomass as energy source (WWF, 2011). Therefore, more areas of tropical forests will be exploited, which increases the potential damage to biodiversity. Logging is one of the main causes of biodiversity loss in tropical forests (Laurance, 2007) due to impacts such as changes in forest structure, disturbance, dislodgement of animals from their habitats, gap opening, and loss of food resources (Lagan et al., 2007).
In the past decades there has been an increase in the number of studies aimed at investigating the effect of logging on the biodiversity of tropical forests, which encompassed several taxa, such as mammals, birds, beetles, and fish (Edwards et al., 2012; Politi et al., 2012). However, the results of most studies have been put into doubt for not being reliable or consistent, mainly due to errors in sampling design, such as pseudoreplication (Ramage et al., 2013). Other problems result from the lack of standardization and from poor baseline information on the study areas, such as management regime and logging period (Laufer et al., 2013).
Specifically for bats, there was little advancement in studies focused on the effect of logging, in spite of the ecological importance of these mammals. Bats are known to be important bioindicators of the status of a given habitat. Their quality as bioindicators results in part from their life history, which is one of the most diversified among mammals (Barclay and Harder, 2003; Jones et al., 2009). In addition, bats are important agents of seed dispersal and pollination, processes that are vital for the reproductive success of plants as well as for the maintenance of tropical forests, may help recover degraded (Fleming and Heithaus, 1981; Fleming and Sosa, 1994; García et al., 2000; Muscarella and Fleming, 2007) and arthropod suppression (Kunz et al., 2011; Williams-Guillen et al., 2008).
Although some studies suggest that the negative effects of logging on bats in the tropical forest with some type of management are minimum (Castro-Arellano et al., 2007, 2009; Clarke et al., 2005a; Presley et al., 2008, 2009) some guilds have been affected, mainly in Latin America. Frequently, after the exploitation there has been an increase in the number of small frugivorous bats in the understory and a decrease in the number of gleaning animalivores, omnivores, and carnivore bats (Clarke et al., 2005b; Peters et al., 2006). However, studies that reported impacts of logging on bats need better planning and sampling design; most of them did not use data on changes in vegetation structure due to logging and they did not test how these changes affect the composition of the bat assemblage (Castro-Arellano et al., 2009; Presley et al., 2008, 2009).
In the present study, we conducted a review of studies on the effects of logging on bats in tropical forests worldwide carried out in the past three decades. We aimed at answering the following questions: What is known about the effects of logging on the bat fauna of tropical forests? What are the gaps of knowledge that can be filled?
Material and methodsWe conducted a literature search for studies published in the past three decades (1988–2012), which assessed the effect of logging on the richness and abundance of bats in tropical forests. We surveyed the databases Web of Science, Google Scholar, and Scopus, using different keyword combinations: “Bats OR Chiroptera”, “Logging”, “Selective logging”, “Timber extraction”, “Tropic”, “Forest”, and “Tropical Forest”.
We used the following criteria to determine whether the reference was suitable for assessment: (1) studies carried out between the latitudes 23 N and 23 S, (2) studies that assessed the effects of logging on bats by comparing areas submitted to some sort of management, such as selective logging, reduced impact logging (RIL), polycyclic selective logging, and continuous selective logging with control areas. We also considered papers with sampling in logging areas, but whose objective was not to assess the effects of selective logging.
In our search we found 118 potential studies, but only 22 fit our criteria. We compiled from those papers the following information: publication year, title, location of the study site (e.g.: geographic coordinates, country), type of management (e.g.: reserve, selective logging, reduced impact logging (RIL), polycyclic selective logging, agriculture, and other types of management), size of the study area (ha) when available, exploitation period (years) since the first cut, volume of timber extracted (m3ha−1), bat sampling methods (mist nets, harp traps, active search in roosts, ultrasound detector), number of individuals collected per site, richness (number of species), trophic guilds (frugivorous, nectarivores, gleaning animalivores, aerial insectivores, omnivores, carnivores, and sanguivores), environmental and disturbance data when available, and sampling effort. As our data were normally distributed (tested with a Shapiro–Wilk test), population's variances were equal (homoscedasticity), and independent (Zar, 1996) we used an analysis of variance (ANOVA) to test for differences in the number of capture methods used and species richness.
We used the Mann–Whitney U-test to compare the most abundant guilds of bats (frugivorous, nectarivores, aerial insectivores and gleaning animalivores) between logged and unlogged forests (Zar, 1996). Only abundance data for bats captured with mist nets were included in this analysis. The abundances were obtained by dividing the number of individuals captures for each trophic guild by the sampling effort for each study with data available. The mist net sampling effort obtained in the studies (Table 1) were standardized in met m2h (sensu Straube and Bianconi, 2002).
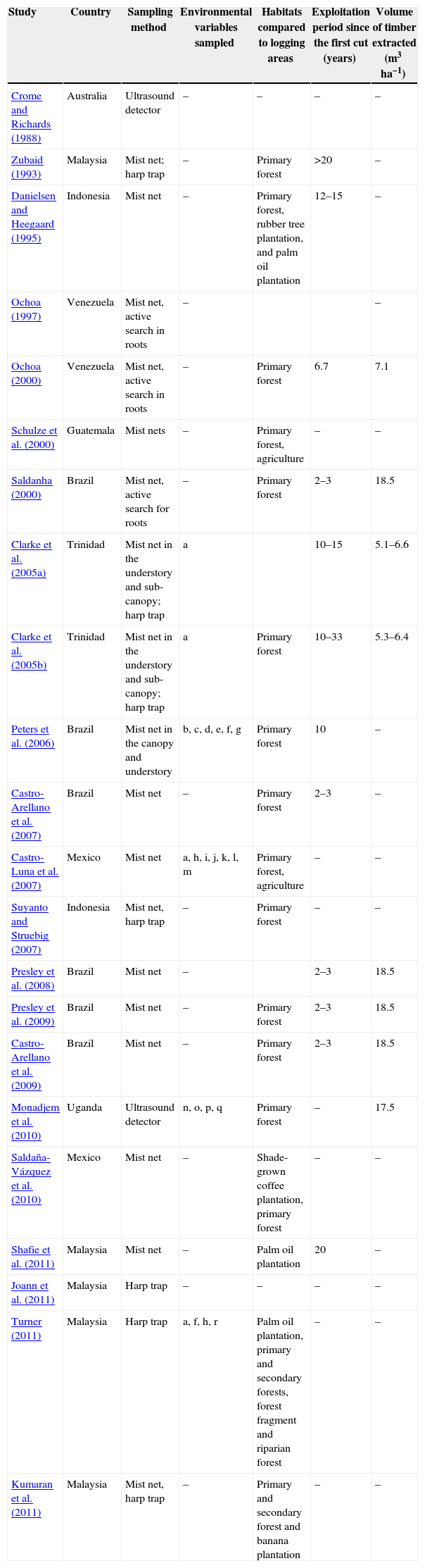
List of studies that investigated the effect of logging on the bat fauna of tropical forests.
| Study | Country | Sampling method | Environmental variables sampled | Habitats compared to logging areas | Exploitation period since the first cut (years) | Volume of timber extracted (m3ha−1) |
|---|---|---|---|---|---|---|
| Crome and Richards (1988) | Australia | Ultrasound detector | – | – | – | – |
| Zubaid (1993) | Malaysia | Mist net; harp trap | – | Primary forest | >20 | – |
| Danielsen and Heegaard (1995) | Indonesia | Mist net | – | Primary forest, rubber tree plantation, and palm oil plantation | 12–15 | – |
| Ochoa (1997) | Venezuela | Mist net, active search in roots | – | – | ||
| Ochoa (2000) | Venezuela | Mist net, active search in roots | – | Primary forest | 6.7 | 7.1 |
| Schulze et al. (2000) | Guatemala | Mist nets | – | Primary forest, agriculture | – | – |
| Saldanha (2000) | Brazil | Mist net, active search for roots | – | Primary forest | 2–3 | 18.5 |
| Clarke et al. (2005a) | Trinidad | Mist net in the understory and sub-canopy; harp trap | a | 10–15 | 5.1–6.6 | |
| Clarke et al. (2005b) | Trinidad | Mist net in the understory and sub-canopy; harp trap | a | Primary forest | 10–33 | 5.3–6.4 |
| Peters et al. (2006) | Brazil | Mist net in the canopy and understory | b, c, d, e, f, g | Primary forest | 10 | – |
| Castro-Arellano et al. (2007) | Brazil | Mist net | – | Primary forest | 2–3 | – |
| Castro-Luna et al. (2007) | Mexico | Mist net | a, h, i, j, k, l, m | Primary forest, agriculture | – | – |
| Suyanto and Struebig (2007) | Indonesia | Mist net, harp trap | – | Primary forest | – | – |
| Presley et al. (2008) | Brazil | Mist net | – | 2–3 | 18.5 | |
| Presley et al. (2009) | Brazil | Mist net | – | Primary forest | 2–3 | 18.5 |
| Castro-Arellano et al. (2009) | Brazil | Mist net | – | Primary forest | 2–3 | 18.5 |
| Monadjem et al. (2010) | Uganda | Ultrasound detector | n, o, p, q | Primary forest | – | 17.5 |
| Saldaña-Vázquez et al. (2010) | Mexico | Mist net | – | Shade-grown coffee plantation, primary forest | – | – |
| Shafie et al. (2011) | Malaysia | Mist net | – | Palm oil plantation | 20 | – |
| Joann et al. (2011) | Malaysia | Harp trap | – | – | – | – |
| Turner (2011) | Malaysia | Harp trap | a, f, h, r | Palm oil plantation, primary and secondary forests, forest fragment and riparian forest | – | – |
| Kumaran et al. (2011) | Malaysia | Mist net, harp trap | – | Primary and secondary forest and banana plantation | – | – |
a, canopy opening (%); b, average diameter of trees; c, basal area; d, coefficient of variation of leaf density in the understory; e, coefficient of variation of leaf density in the canopy; f, leaf density in the understory (<5m); g, leaf density in the canopy (5–40m); h, height of the canopy (m); i, herbaceous cover (%), j, woody plants cover (%), k, plant diversity (H′); l, distance from the forest edge (m); m, distance from caves (m); n, vegetation cover (0–5) in the understory; o, vegetation cover (0–5) in the sub-canopy; p, vegetation cover (0–5) in the canopy; q, temperature (°C), r, number of water bodies.
We used ArcGis 9.3 (ESRI, 2009) to draw the distribution map of studies (Fig. 1). To generate the map, we used the geographic coordinates provided in each study. When this information was not available, we used Google Earth to obtain approximate coordinates of the study areas, using as reference characteristics that could be easily identified in the images, such as rivers, protected areas, cities, and roads, which were mentioned in the papers.
Results
We found 22 publications about the effect of logging on bats in tropical forests, which were published in the past three decades. Most studies were carried out in the countries of Latin America (Brazil=27%, Mexico=9%, Venezuela=9%, Trinidad=9%, Guatemala=5%) and Southeast Asia (Malaysia=23%, Indonesia=9%). We found only one study for the tropical forests of Africa and one for the tropical forests of Oceania (Table 1 and Fig. 1). In a temporal analysis considering the past three decades there was an increase in the number of publications. The period from 2005 to 2010 was the most productive (Fig. 2), whereas from 2002 to 2004 we did not find publications on the topic (Table 1).

Most studies (81.8%) only compared richness and abundance between logged and unlogged areas. In Latin America, most studies (40%) compared logged areas that presented some management regime, mainly reduced impact logging, with unexploited areas, as observed in studies carried out in the Brazilian Amazon (Castro-Arellano et al., 2007, 2009; Presley et al., 2008, 2009; Saldanha, 2000). Only one study in Trinidad (Clarke et al., 2005a) assessed the effect of two logging systems on the bat assemblage: continuous logging, which is a system of continuous harvest that has no ecological criteria, and periodic blocks (PB), which is a polycyclic system based on ecological criteria aimed at biodiversity maintenance. In Southeast Asia, in spite of intensive logging activity, only two studies investigated the effect of logging on bats (Danielsen and Heegaard, 1995; Turner, 2011). The other studies carried out in Southeast Asia only mentioned that the study was conducted in logging sites without making any analysis on the topic.
Few studies used environmental and disturbance variables, such as the alterations suffered in the vegetation structure, to interpret the effect of logging on the bat fauna, which might lead to more precise conclusions. Out of 22 studies found, only four (18%) used this kind of variables (Table 1 and Fig. 3). Two studies only measured canopy opening, but did not report how this variable could affect the bat fauna in Trinidad (Clarke et al., 2005a, 2005b).
; EP, exploitation period since the first cut (years); AB, abundance of bats in studies in tropical forests published from 1985 to 2012. In red: percent of studies which reported data. In blue: percentage of studies which did not reported data.")
Proportion of number of studies according to data presented. EV, environmental variables sampled; VE, volume of timber extracted (m3ha−1); EP, exploitation period since the first cut (years); AB, abundance of bats in studies in tropical forests published from 1985 to 2012. In red: percent of studies which reported data. In blue: percentage of studies which did not reported data.
Only eight studies (36%) provided data on the volume of timber exploited per hectare (m3ha−1), with values ranging from 5.1 to 18.5m3ha−1 (Table 1 and Fig. 3). Two studies informed the number of trees removed per hectare (Danielsen and Heegaard, 1995; Peters et al., 2006). The period of exploitation (years) since the first cut was provided for more than 50% of the studies (Table 1 and Fig. 3). The size of the study area (ha) was provided for 68% of the studies. Sixteen studies (72%) informed data on bat abundance (Fig. 3).
Half of the studies (11) used mist nets. Two studies used ultrasound detectors, two studies used harp traps, and seven studies used two or more methods combined (Table 1). Considering the number of species captured (richness) with each sampling method, there was a significant difference (F=7.35; p<0.012). In studies that used one single sampling method, the number of species captured ranged from three to 35. With two methods the range was broader: from two to 78 species. We observed the smallest range in studies that used three methods: from 18 to 30 species. We found different ways to describe the sampling effort. For example, Clarke et al. (2005a) described it as netshours, Presley et al. (2009) as netsmetershours, and Kumaran et al. (2011) as nets×night. This lack of standardization hinders comparisons between studies.
Considering the number of individuals captured by trophic guild in studies that used mist nets to capture bats, we found a trend of increase in abundance of frugivorous bats and a decrease of gleaning animalivores bats in logged forests. However, this trend was only marginally significant for gleaning animalivores (U=95.5, p=0.050), and was not significant for nectarivores (U=80.5, p=0.520), frugivorous (U=146.0, p=0.245), and aerial insectivores (U=64.0, p=0.207) when comparing logged and unlogged forests (Fig. 4).
Discussion
Bats have been overlooked in assessments of the effect of logging on tropical forests worldwide. However, we found that there was an increase in studies with this group in the past decades. Among the few studies found in the literature, most were conducted in the Neotropics, in terra firme forests and in areas with some regime of management with very low timber extraction (Castro-Arellano et al., 2007, 2009; Clarke et al., 2005a, 2005b; Ochoa, 2000; Peters et al., 2006; Presley et al., 2008, 2009; Saldanha, 2000). The tropical forests of Africa and Oceania were the ones with least number of studies. This trend created a deficit in our understanding of bat responses to logging, considering that the fragmentation caused by logging may alter the trophic organization of communities (Hill et al., 2011).
We found that these studies assessed only differences in richness, abundance, and food guilds between logged and unlogged areas. For instance, they did not include data on environmental changes caused by logging, such as changes in forest structure. One of the few studies that considered changes in vegetation structure was carried out in the Brazilian Amazon (Peters et al., 2006), where the effect of selective logging in areas exploited for ten years was analyzed. The authors observed that changes in canopy opening and leaf density in the understory caused by logging lead to changes in the composition and trophic structure of the bat fauna. This may suggest that selective logging does not have a high conservation value. However, the above-mentioned study has methodological flaws, such as pseudoreplication, which may lead to exaggerated results in studies on the effects of selective logging on biodiversity (Ramage et al., 2013).
Studies on the effects of logging on bats have used mist nets as the main sampling method, probably because mist nets are relatively accessible, cheap, easy to transport in the field, and quite efficient to capture bats (Kunz et al., 2011). However, in the Neotropics mist netting favors the capture of phyllostomids, which are endemic to this region, which leaves other bat families undersampled in studies on the effect of logging in tropical forests (Presley et al., 2008, 2009). The use of complementary methods, such as active search for roosts, an efficient and not very expensive method, would solve this methodological problem (Ochoa, 2000). Ultrasound detectors (Cunto and Bernard, 2012) could also be used for sampling bats that forage in the canopy, such as vespertilionids. In addition, our review of the literature of the past three decades pointed to an increase in the number of methods used from 2004 to 2006, which was not maintained in the subsequent years.
In most studies there was a consensus that logging has a positive effect on frugivorous and nectarivores bats, and a negative effect on gleaning animalivores bats, which are bats that feed on invertebrates and small vertebrates picked from the substrate (Giannini and Kalko, 2004). The positive effect on frugivorous and nectarivores bats probably results from the use of food provided by pioneer plants that flourish in response to changes caused by logging (Clarke et al., 2005b; Fleming, 1988; Presley et al., 2008; Soriano and Ochoa, 2001). Conversely, the negative effect on gleaning animalivores bats is probably related to a decrease in the abundance of prey and a decrease in the number of roosting sites (Presley et al., 2008; Soriano and Ochoa, 2001). In addition, in some cases these bats depend on specific kinds of roosts that are destroyed by logging (Kalko, 1998).
Although we only found a marginally significant decrease of gleaning animalivores bats in logged forests, our data suggests that there was an increase in the abundance of frugivorous bats in logged forest. The positive effect on frugivorous bats probably results from the use of food resource provided by pioneer plants that flourish in response to changes caused by logging (Clarke et al., 2005b; Fleming, 1988; Presley et al., 2008; Soriano and Ochoa, 2001). Conversely, the negative effect on gleaning animalivores bats which are bats that feed on invertebrates and small vertebrates picked from the substrate is probably related to a decrease in the abundance of prey and a decrease in the number of roosting sites (Presley et al., 2008; Soriano and Ochoa, 2001). In addition, in some cases these bats depend on specific kinds of roosts that are destroyed by logging (Kalko, 1998). Our data also showed a large variation in the abundance of frugivorous bats in logged forests, which probably diluted the logging effect on this trophic group. Thus, a more detailed analysis, considering an overview on the species level would probably show that some species may respond more positively than others withing this sepcific guild.
Thus, we conclude that the effects of logging on bats in tropical forests require more research for better comprehension. However, our results suggest that for frugivorous bats the effects tends to be positive while for gleaning animalivores the effects tends to be negative.
In short, in order to understand the responses of bats to logging in tropical forests, considering the ecological importance of these animals (Kunz et al., 2011), future studies should invest in better planning. We suggest some guidelines for their improvement, such as the standardization of sampling design, in order to avoid pseudoreplication, which may undermine the results and conclusions. Complementary sampling methods should be used, such as mist netting and active search, mist netting and ultrasound detectors, or mist netting, active search, and ultrasound detectors. We also suggest the inclusion of environmental and disturbance variables in the analysis of changes that affect the richness and abundance of bat species or guilds. Finally, for futures studies we suggest more attention to poorly studied regions with intensive logging activity, such as the Amazon and the tropical forests of Africa.
Conflicts of interestThe authors declare no conflicts of interest.

