



Land degradation is a complex concept that integrates different aspects, including changes in soil conditions, biodiversity, productivity and socio-economic implications, compared to a reference state. We propose a new conceptual framework to analyze degradation stages and restoration thresholds in species-rich natural grasslands. The framework integrates different degradation stages with their respective thresholds and describes key processes of land-use change that lead to certain stages and thresholds. Specifically, we discuss two scenarios of grassland degradation, i.e. unsuitable grassland management and complete change of land use, sometimes followed by spontaneous recovery. We illustrate the framework with the case of south Brazilian grasslands, which are rich in biodiversity, but suffer from a series of degradation processes and are poorly considered from a conservation perspective. The conceptual framework can be applied by studies on degradation and restorability of tropical and subtropical grasslands after changes in management or transition to other land use; it will facilitate decisions on alternative management and conservation.
Land-use change and degradation of natural ecosystems are principal causes for losses of biodiversity and ecosystem functions (Sala et al., 2000). Concrete numbers on land degradation are often only available for the complete loss of natural ecosystems, for example after conversion to more productive systems such as arable fields or tree plantations (for Brazil e.g. IBGE, 2012). However, degradation also includes less marked changes in structure, composition or ecological processes within the ecosystem. Therefore, degradation needs to be analyzed at different spatial scales, from a local focus on specific degradation processes to landscape and regional scales, and using diverse methods including remote sensing, plot-based measurements, experiments, expert knowledge and assessment of stakeholder experience (Reed et al., 2011). To provide a better understanding of the dynamics of degraded ecosystems and to facilitate mitigation of degradation processes and restoration, the concept of thresholds between alternative stages has been developed. While the framework suggested by Briske et al. (2006) focuses on definition and description of thresholds, Hobbs et al. (2009) described ‘historical’, ‘hybrid’ and ‘novel’ stages and identified ‘restoration thresholds’ when ecological, cultural or technical obstacles prevent a system to return to its original state. However, a synthesis of these approaches is missing so far.
Grasslands are among the ecosystems with highest species richness in the world (Wilson et al., 2012), and they provide a wide range of ecosystem services. Grasslands play an important role within the global carbon cycle, as 90% of their biomass is belowground, accumulation rates are high, and decomposition of organic material slow (Gibson, 2009). As main forage resource for livestock, grasslands are important for human well-being in many regions. They facilitate infiltration of water into the soil and thus to the maintenance of hydrological cycles. Finally, grasslands contribute to the landscape beauty of many regions. Thus, they are multi-functional systems but at the same time subjected to unsustainable use and conflicting interests.
Conceptual frameworks of grassland degradation have been developed specifically for rangelands, i.e. grazed systems, where stocking rates often are inadequate (e.g. Bestelmeyer, 2006). At the same time, large areas of grasslands are endangered due to land-use changes or have already been lost (Sala et al., 2000), especially in tropical and subtropical regions (Bond and Parr, 2010). Elsewhere, cropland on sites that originally had been grassland is abandoned for economic reasons, and grassland re-assembles spontaneously under re-introduction of moderate grazing management (Hölzel et al., 2002; Öster et al., 2009). This latter scenario, to our knowledge, has not yet been integrated in grassland or rangeland degradation models.
In this paper, we present a conceptual framework of grassland degradation that for the first time integrates different degradation stages and two types of recovery thresholds, including the most important processes related to land-use history causing stage transitions. We illustrate the framework with studies from grasslands in Rio Grande do Sul (hereafter: RS) State, southern Brazil, where biodiversity is well studied, but where conservation of grassland has been neglected, and degradation processes are poorly studied (Overbeck et al., 2007, 2013). Finally, we point out how current limitations regarding knowledge on degradation processes and conservation state of grasslands should guide future research.
Conceptual framework of grassland degradation and restorationThe starting point of our framework is the use of traditionally managed grasslands as reference systems. This reference stage is comprised by grasslands composed of native species, with a specific and high biodiversity and considerable conservation value, usually due to extensive grazing and associated management practices (e.g. fire, mowing). The importance of management for maintenance of these systems and their diversity has been shown before (e.g. Overbeck et al., 2007; Pillar and Vélez, 2010 for southern Brazil).
As in Hobbs et al. (2009), changes of ecosystem properties can be displayed as biotic and abiotic changes at the local scale along two axes (Fig. 1). As biotic changes we consider deviation in species composition, vegetation structure (vertical and horizontal structural variables), basic ecological processes (e.g. pollination, seed dispersal), resulting from altered management, species introduction or conversion of land use. This includes alien species invasion. As abiotic changes we understand alteration in soil chemical and physical properties, as caused by fertilization or soil cultivation. The applicability of the framework is directly related to the quantification of these factors in case studies and the establishment of the thresholds according to changes in biotic and abiotic conditions in grasslands. It allows us to identify the stage of degradation of a given area and its potential for restoration. Changes in grassland management will cause properties of the system to change gradually, resulting in a decrease in resilience, but with resumption of the historical management the original properties might be reached again in many cases, i.e. a self-recovery threshold is not crossed (Fig. 1a, Scenario 1). If the grassland is converted to other land use, this will lead to an almost entire change of the original properties, e.g. complete loss of the aboveground plant community (Fig. 1b, Scenario 2), while abandonment of these land uses initiates re-colonization from the regional species pool (Fig. 1c). This process is of particular importance in regions of the world where grassland restoration techniques have not been developed. In this context, it needs to be recognized that spontaneous recovery may often mean colonization of non-typical or alien species, i.e. trajectories of community development that not necessarily lead to the desired state.
 properties of the reference grassland are moderately altered if grassland management is changed, but modification is reversible; (b) after conversion to other land uses, the properties of the reference grassland are radically changed and ecosystem resilience is lost; (c) after abandonment some properties might recover (‘self-recovery threshold’) or be restorable (‘restoration threshold’); and (d) integration of the various scenarios.")
Conceptual framework on degradation and restoration of species-rich natural grasslands: (a) properties of the reference grassland are moderately altered if grassland management is changed, but modification is reversible; (b) after conversion to other land uses, the properties of the reference grassland are radically changed and ecosystem resilience is lost; (c) after abandonment some properties might recover (‘self-recovery threshold’) or be restorable (‘restoration threshold’); and (d) integration of the various scenarios.
In the framework, the distinction between two types of thresholds is important: (1) a self-recovery threshold that describes until which point a recovery without additional technical measures is possible, and (2) a restoration threshold that identifies until which point an area can be restored with technical assistance (e.g. modification of soil features, control of undesired species, introduction of desired species).
South Brazilian native grasslands: origin and developmentIn RS, grasslands occur in the highlands in the north of the state, where they form mosaics with Araucaria forest, and in its southern half, in the Pampa biome, where they dominate the landscape (Overbeck et al., 2007), continuing in Uruguay and Argentina (Fig. 2). South Brazilian grasslands are particularly rich in plant species, with about 2200 grassland plant taxa known only for RS (Boldrini, 2009).

Grasslands in the region are primary. They are relicts from cooler and drier periods, and were affected by forest expansion since approximately 5000 years BP, favoured by warmer and more humid climate, with increasing rates since 1500 years BP (Behling, 2002). After the extinction of large herbivores (Lima-Ribeiro and Diniz-Filho, 2013), these grasslands had been maintained by anthropogenic fires and by grazing of small mammals (Cione et al., 2003; Behling and Pillar, 2007), and since the 17th century by introduced livestock. Today, beef production is an important economic activity in the region, with native plant species constituting most grassland vegetation. Available data indicate that plant diversity and forage production are highest under intermediate levels of grazing and intermediate fire frequency (Overbeck et al., 2005; Nabinger et al., 2009). Management thus can be considered essential for preservation of grassland biodiversity, as observed in many ‘old-growth grasslands’ (Veldman et al., 2015). However, transformation rates are high: between 1986 and 2002, grassland areas suffered losses of 16%, which corresponds to a loss rate of 1000km2 per year (Cordeiro and Hasenack, 2009).
Land-use change in the grassland region according to remote sensingEvaluation of LANDSAT data shows that ca. 60% (104,553km2) of former grassland area in southern Brazil had been destroyed by 2002 (Fig. 3), mostly due to the conversion to arable fields or alien tree plantations. Losses of native grassland have not been uniform in space, but reflect soil properties and topographic constraints. In the Central Western Plateau region, native grasslands were nearly completely transformed into cropland, mostly for soybean. The coastal region has seen high rates of transformation, principally to rice and pine plantations. In the Northeastern Plateau, where soils are shallower, tree plantations and arable fields are the main causes of grassland losses. Here, land-use change has increased considerably within the past decade.
. Grassland areas with clearly visible signs of former land-use change (e.g. use as agricultural field) are considered as degraded, i.e. reflect past land-use change. Original grassland areas that have been completely transformed to other uses and not recovered to grassland comprise the category ‘converted’. Regions in white are those where natural vegetation cover is forest.")
Distribution of grassland remnants and degraded grassland in RS State. The map is based on Landsat ETM+ images (spatial resolution: 30m). Grassland areas with clearly visible signs of former land-use change (e.g. use as agricultural field) are considered as degraded, i.e. reflect past land-use change. Original grassland areas that have been completely transformed to other uses and not recovered to grassland comprise the category ‘converted’. Regions in white are those where natural vegetation cover is forest.
In the different regions of the state, 5–17% of the grassland area is classified as ‘degraded’, i.e. remote sensing data indicate former agricultural activity (e.g. tillage lines). Returning these areas to high nature conservation status might compensate for some of the ongoing losses through grassland conversion. However, compositional characteristics of vegetation itself (e.g. presence of alien species) cannot be observed by remote sensing data at this scale. In the Central Depression and in the Southwestern Grasslands of RS, for instance, a considerable proportion of the remaining grasslands mapped as ‘conserved’ actually have been degraded by alien forage species, which were deliberately seeded in some areas and colonized others, or by other processes. On-site ground studies are indispensable to verify the level of degradation and existence of self-recovery and restoration thresholds; remote sensing data alone can only give a limited picture of grassland degradation.
Scenario 1: Degradation of grasslands after changes in management, and potential for self-recoveryGrazingMost studies evaluating effects of different grazing intensities in RS focus on effects on forage or beef production (e.g. Maraschin and Corrêa, 1994; Moojen and Maraschin, 2002; Pinto et al., 2008), and only few analyze effects on species composition (Boldrini and Eggers, 1997; Soares et al., 2011) or soil properties (Bertol et al., 1998). Usually, grazed grasslands with moderate grazing intensity are formed by mosaics of intensively grazed patches dominated by prostrate grasses (e.g. Axonopus affinis Chase, Paspalum notatum Flüggé), and less grazed patches dominated by tussock grasses, small shrubs or other species less attractive for grazing animals (Boldrini and Eggers, 1997; Diaz et al., 2007). This heterogeneity of the vegetation leads to structural complexity and diversity.
If grazing is excluded – until now, a common practice in conservation units of Rio Grande do Sul – grassland structure quickly changes: tall tussock grasses, e.g. Andropogon and Sorghastrum spp. (Boldrini and Eggers, 1997), become dominant, litter accumulates and microclimate at soil surface changes (Pallarés et al., 2005), drastically reducing plant species richness (Overbeck et al., 2005). In northeastern RS, the encroachment of shrubs of the genus Baccharis (principally, B. uncinella DC) and a slow invasion of forest pioneer species have been observed after abandonment (Oliveira and Pillar, 2004) despite the accumulation of grass biomass that may hinder fast recruitment of woody pioneers.
Overgrazing, on the other hand, can lead to the replacement of productive forage plants by species with lower forage quality, resulting in increasing cover of ruderal species and bare soil, while the contribution of highly nutritional C3 grasses decreases (Pallarés et al., 2005). Ecosystem functions like water infiltration can be negatively affected as soil bulk density increases (Bertol et al., 1998). Either situation, when grazing is excluded or when the grassland is overgrazed, may be considered degraded due to changes in biotic and abiotic characteristics and the reduction of ecosystem resilience.
Introduction of alien C3 and C4 species and fertilizationOverseeding of natural grasslands with introduced alien species, often combined with fertilization and liming, aims to increase forage quality and quantity especially in winter (Nabinger et al., 2000). Although many native species have high productivity and nutritive potential, they are not available on the seed market, and introduced species are used instead; common species are Lolium multiflorum Lam., along with some European Fabaceae, e.g. Trifolium repens L. It has been shown that forage yield increases linearly with nitrogen addition (Brambilla et al., 2012) and promotes the increase of animal live weight gain per area, but also leads to marked changes in the floristic composition (Pallarés et al., 2005; Brambilla et al., 2012).
Introduced alien grassland species may become a serious problem when they spread to natural ecosystems. Large-scale invasion of these European-origin species that are used as forage, however, has not been reported yet, in contrast to some other species from temperate climates (e.g. Ulex europaeus L.) and, principally, some C4 grass species of African origin. For example, Eragrostis plana Nees had invaded up to 20% of Rio Grande do Sul's grassland area by the year 2008, causing severe reduction in forage quality and native plant diversity (Medeiros et al., 2004). Medeiros and Ferreira (2011) succeeded in at least partial suppression of populations of the invasive species by a combination of soil cultivation and seeding of both native and non-native forage species.
FireFire has shaped the southern Brazilian grasslands during the past millennia (Behling and Pillar, 2007). The use of burns as management tools for livestock production, traditionally applied in the highland grasslands, is controversial due to concerns regarding possible negative impacts, and thus fire had been prohibited by state legislation until recently. Regular fires select for different species groups compared to grazing (tussock vs. prostrated grasses, respectively), and selectively affect some species groups (e.g. C3 grasses when burns occur in winter), but do not seem to cause reductions in grassland diversity (Overbeck et al., 2005). Fidelis et al. (2012) showed higher species richness in frequently burned grassland plots in comparison to sites where burning and grazing had been excluded for some years. In fact, a large part of the grassland species is adapted to fire and can resprout from underground storage organs (Fidelis et al., 2014). Invertebrate communities and processes determined by their activity, e.g. decomposition, return to pre-fire values after relatively short periods, at least under patchy fire at fine scale (Podgaiski et al., 2013, 2014). Exclusion of fire in ungrazed areas, a common practice in conservation units, leads to the accumulation of dead biomass and the risk of high-intensity fires increases.
Potential for self-recovery and restoration after management-induced degradationPotential for self-recovery should be high if the species present are members of the characteristic species pool of the region and changes in composition and structure were provoked mainly by changes in management. When soil conditions were changed, e.g. by liming or fertilization, self-recovery may be more difficult or even impossible, because introduced, sometimes invasive, species with high competitive abilities may be able to maintain high cover. Current research suggests that prescribed fire could be a conservation tool in areas where grazing is not feasible (Overbeck et al., 2005; Fidelis and Blanco, 2014), e.g. in conservation units. Thus, intensity and frequency of management are key factors for grassland recovery after any kind of degradation (Winter et al., 2012).
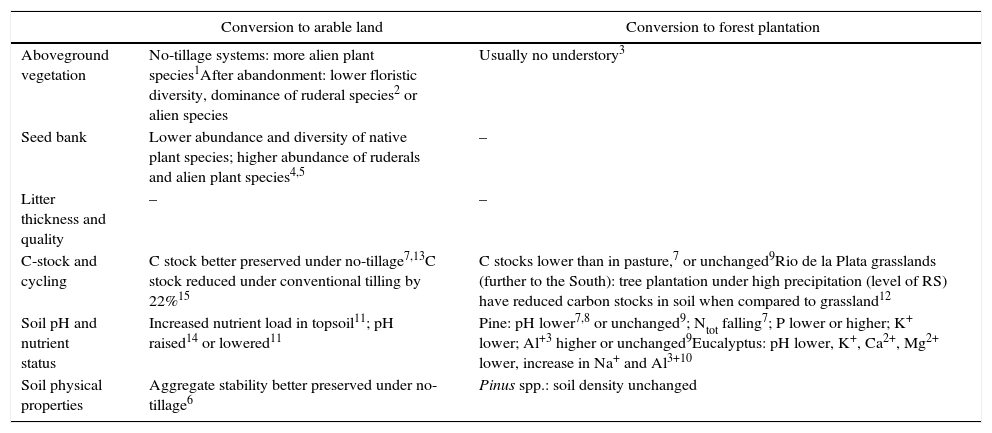
Scenario 2: Degradation and potential for self-recovery after complete conversion of grasslandFor southern Brazil, some studies on ecosystem properties and ecological processes under different types of land use in former grassland areas are available. Table 1 synthesizes the available data, considering variables of importance for regeneration or restoration after the end of intensive land use (e.g. seed bank) or that may persist in a changed condition over long periods of time.
Review of studies on the effects of land-use change on ecosystem processes in grasslands of southern Brazil (RS, Santa Catarina, Paraná). All trends in comparison to reference grasslands (–, no studies available).
| Conversion to arable land | Conversion to forest plantation | |
|---|---|---|
| Aboveground vegetation | No-tillage systems: more alien plant species1After abandonment: lower floristic diversity, dominance of ruderal species2 or alien species | Usually no understory3 |
| Seed bank | Lower abundance and diversity of native plant species; higher abundance of ruderals and alien plant species4,5 | – |
| Litter thickness and quality | – | – |
| C-stock and cycling | C stock better preserved under no-tillage7,13C stock reduced under conventional tilling by 22%15 | C stocks lower than in pasture,7 or unchanged9Rio de la Plata grasslands (further to the South): tree plantation under high precipitation (level of RS) have reduced carbon stocks in soil when compared to grassland12 |
| Soil pH and nutrient status | Increased nutrient load in topsoil11; pH raised14 or lowered11 | Pine: pH lower7,8 or unchanged9; Ntot falling7; P lower or higher; K+ lower; Al+3 higher or unchanged9Eucalyptus: pH lower, K+, Ca2+, Mg2+ lower, increase in Na+ and Al3+10 |
| Soil physical properties | Aggregate stability better preserved under no-tillage6 | Pinus spp.: soil density unchanged |
1Favreto et al. (2007); 2Boldrini and Eggers (1997); 3Souza et al. (2013); 4Favreto and Medeiros (2006); 5Maia et al. (2008); 6Bertol et al. (2004); 7Wiesmeier et al. (2009); 8Schumacher et al. (2008); 9Mafra et al. (2008); 10Céspedes-Payret et al. (2012); 11Rheinheimer et al. (1998); 12Berthrong et al. (2012); 13Pillar et al. (2012); 14Almeida et al. (2005); and 15Diekow et al. (2005).
Studies on the seed bank of arable fields on former grasslands in southern Brazil show that the number of grassland species decreases with management intensity, giving place to native or alien ruderal species (Favreto et al., 2007), thus reducing recovery potential of grassland. These results are in line with studies from other grassland ecosystems around the world that show higher abundance of weed species in the seed bank after agricultural use (Hutchings and Booth, 1996; Kiehl and Pfadenhauer, 2007). As vegetative recovery is the principal regeneration strategy of South Brazilian grasslands after disturbance (Fidelis et al., 2009), seed input from external sources as well as abiotic conditions should be limiting for recovery of the former grassland community: the bud bank likely does not persist through periods of intensive agricultural use.
Grassland conversion also results in changes of soil properties. Arable land use increases nutrient levels of the soil (Rheinheimer et al., 1998; Perin et al., 2003), leading to different trajectories of vegetation recovery. It is well known that large quantities of carbon stored in grasslands may be rapidly transferred to the atmosphere and lost when the grassland is plowed and converted to arable fields (Sala and Paruelo, 1997; Pillar et al., 2012). In a worldwide meta-analysis of carbon changes due to land-use changes, Guo and Gifford (2002) showed that a conversion of grasslands to crop rotation leads to a loss of 60% of belowground carbon. For southern Brazil, a decrease in C-stock in soils under conventional-tilling has been shown, with magnitude depending on management intensity; no-tillage systems result in much lower losses of C in soils (Bertol et al., 2004; Diekow et al., 2005).
Tree plantationsBy 2009, 6000km2 (10%) of grasslands in RS were converted to plantations of pines, eucalypts or acacias (Gautreau and Velez, 2011). Native grassland vegetation composition is drastically affected under tree plantations, even with reduced soil disturbance for tree planting (Pillar et al., 2002). Gonçalves et al. (2008) found relatively low species richness and dominance of a few ruderals and some alien species like the grass Melinis minutiflora P. Beauv. in the soil seed bank under tree plantations in the Central Brazilian cerrado. Likewise, and in analogy to former agricultural fields, we can expect a low contribution of the seed bank in vegetation recovery.
Studies on effects of tree plantations on grassland soils give variable results (Table 1). Guo and Gifford (2002) stated that the conversion to plantations leads to a significant reduction of soil C stocks when coniferous species were used, while the effect with broadleaf species like eucalypts was not significant. For RS, Wiesmeier et al. (2009) found lower C stocks under pine plantations, while Mafra et al. (2008) could not show any changes. A growing number of literature examining potential for carbon sequestration in plantations is available, with changes based on the shift from belowground biomass dominance (grassland) to aboveground biomass with litter accumulation (plantation) (Guo et al., 2008). It has been shown that this potential strongly depends on soil types (Zinn et al., 2002), and might not be true for regions with high precipitation like RS, for which a decrease in soil carbon was observed (Guo and Gifford, 2002; Berthrong et al., 2012).
Potential for self-recovery and recovery thresholds after land-use changeObservational data indicate that the type of vegetation that develops after logging and abandonment differs considerably from that of reference grassland, and that species introduction likely is important if the objective is to restore grasslands. Zaloumis and Bond (2011) showed that species composition of grasslands established after logging of Pinus elliottii plantations in South Africa was markedly changed in comparison to reference grasslands. To what extent spontaneous regeneration will allow for return to pre-disturbance conditions is currently unknown and likely will depend on the time period of other land use (and with this, to what extent seeds or underground organs of target species remain in the ground), intensity of modifications of the site conditions (e.g. fertilization), and the landscape context (i.e. propagule sources). Both biotic and abiotic thresholds for recovery may exist, making active restoration necessary. As grasslands depend on management, intensity and/or frequency of grazing or fire will probably be crucial for the restoration process.
Discussion and conclusionsLand degradation studied on a regional scale often only considers conversion or complete losses of natural ecosystems, with limits to detect e.g. compositional changes, while at the local scale, finer effects of land management have to be considered. Both perspectives are necessary for an improved assessment of degradation and potential restoration, and differences between the two types of degradation likely imply different perceptions regarding degradation, conservation or restoration.
For our study system, conclusive evidence is available that management is necessary for maintenance of diverse and productive grasslands in this region (Overbeck et al., 2007), and biodiversity conservation and livestock production can be considered as complementary management goals, allowing for sustainable use (e.g. Nabinger et al., 2009). Fire and grazing are selective forces that cause changes in grassland composition and structure, but their effects depend on frequency and intensity – both can contribute to conservation of biodiversity and productivity, but they can also be detrimental when frequency or intensity are too high or too low. A systematic and large-scale quantification of effects of different management types (especially intensification, Scenario 1) on different properties of the grasslands in the region is still missing, making it difficult to define degradation more precisely. Furthermore, the necessity of an integrated perspective of biodiversity conservation and sustainable use still is not widely recognized in the debate on conservation strategies (Overbeck et al., 2007; Pillar and Vélez, 2010).
Even though a considerable proportion of natural grasslands in southern Brazil have been converted to other land use (Scenario 2), concerns on potential restoration of these areas have been raised only recently (Overbeck et al., 2013), and are affecting the agenda of conservation authorities. Once supported by additional empirical data, our conceptual framework can support decision making and priority setting in nature conservation by identifying whether costly restoration measures will be necessary or adaptation in management could be sufficient for self-recovery. In this, it is important to recognize that not only biotic and abiotic characteristics are covered by the framework, but that these can also be interpreted in terms of ecosystem functions and services (e.g. carbon sequestration, forage production).
Bestelmeyer (2006) points out problems and risks associated with threshold models: for instance, no single predictive thresholds – which would greatly facilitate management decisions – should be expected to exist, and parameters may reflect measurability, and not long-term degradation processes. Threshold models may become ‘insidious’ (Bestelmeyer, 2006), if they lead to the belief that certain areas are not restorable anymore, because some original features of the system cannot be recovered. This, however, is not a consequence of the model per se, but of a failure of recognizing the full range of features, processes and services of any type of ecosystem. The current debate on novel ecosystems (Hobbs et al., 2013) is centering exactly on the question of how to deal with systems that cannot be brought back to their original state. A conceptual framework of degradation and restoration based on a variety of biotic and abiotic variables (and their interactions, e.g. soil–plant feedbacks, e.g. Suding et al., 2013) has the potential to include different functions and services, and can contribute to a broader understanding of landscapes as multifunctional systems.
Young (2000) suggested that ecological restoration is the future of biological conservation. This means that degraded systems need to become a research focus, even in megadiverse countries where knowledge on biodiversity and functioning of natural ecosystems still has priorities. Conceptual frameworks such as the one presented here can support the study of degradation, conservation and restoration, in southern Brazil and elsewhere. In order to assess conservation or degradation state and restoration potential of degraded systems and to define the recovery and restoration thresholds, it is necessary to collect data on the full gradient from conserved to degraded systems, on a regional scale. For this, we need to develop rapid assessment methods of different parameters of the system, including abiotic and biotic variables as well as measures for ecosystem functions, as proposed by Meyer et al. (2015).
Conflicts of interestThe authors declare no conflicts of interest.
This contribution results from a CNPq-DFG cooperation project (490069/2011-8 and KO1741/2-1, respectively). CK receives support by the Evangelisches Studienwerk Villigst. BA by CAPES. EV, IB and VP receive support from CNPq.

