





Improving management practices in coffee agroecosystems to achieve sustainability is required to counter the current anthropogenic pressures on biodiversity. Identifying the specific habitat characteristics that determine the occupancy of insectivorous birds in coffee farms can represent an effective tool to improve management practices and bird conservation with minimal effort from coffee growers, thus fostering sustainability. To explore this, we modeled the occupancy of six functional groups of birds associated with the insect pest control in response to seven habitat characteristics of interest for bird conservation and coffee habitat management. In general, our results indicate that occupancy probability was correlated with habitat characteristics describing increased vegetation structure across the landscape. Epiphytes strongly influenced all studied functional groups. Most groups were also positively related with herb and shrub cover. However, contrary to our predictions, coffee dominance increased the occupancy of most insectivores, possibly due to the diverse vegetation structure and organic management practices of coffee farms. To provide management recommendations, we identified critical values for each habitat characteristic where the occupancy of most functional groups was benefited. These management recommendations sought to benefit species associated with key ecosystem services, thus possibly improving both conservation and production outcomes.

Improved agroecosystem practices that optimize the provisioning of ecosystem goods and services are essential for achieving agricultural sustainability and biodiversity conservation in the Anthropocene (Kremen and Merenlender, 2018; Mamabolo et al., 2020). Traditional polyculture and rustic shaded coffee agroforestry systems (Moguel and Toledo, 1999) promote biodiversity by maintaining higher levels of species diversity and complementarity between farms and forests (Haggar et al., 2019; Manson et al., 2008) than more intensively managed farms. Intensification of farming practices can also be detrimental to the provisioning of ecosystem services (Cerda et al., 2017; De Beenhouwer et al., 2013). Given the current threats to shade coffee (Harvey et al., 2021) and its importance for tropical conservation efforts and human welfare, additional studies that help identify more sustainable management practices that enhance biodiversity and productivity are warranted.
Since changes in coffee management practices and vegetation structure associated with the intensification of production (Cerda et al., 2017) can affect bird communities (Rodrigues et al., 2018) and their phylogenetic diversity (Frishkoff et al., 2014), this taxa is ideal for assessing the sustainability of coffee management practices (Gregory and Strien, 2010). Birds also provide a host of on-farm services including pollination, seed dispersal, and pest control (Chain-Guadarrama et al., 2019; Michel et al., 2020). This latter service is particularly relevant to mitigating losses from pests and diseases in farms and even enhancing other ecosystem services like bee pollination, increasing coffee productivity (Díaz-Siefer et al., 2022; Martínez-Salinas et al., 2022). Rustic and diversified shade coffee polycultures can increase the abundance, total diversity, and functional richness of insectivorous birds (Bakermans et al., 2012; Martínez-Salinas et al., 2016; Philpott and Bichier, 2011). Therefore identifying habitat characteristics that influence the distribution and abundance of this functional guild may help generate management recommendations that optimize conservation and productivity in these agroecosystems (Sekercioglu, 2012). Moreover, management-induced changes in vegetation structure of agroforestry systems (Jedlicka et al., 2021) are considered a key determinant of bird habitat selection (Cody, 1981; Philpott and Bichier, 2011). Generally, heterogeneity of habitat characteristics in agricultural systems can promote bird diversity (Anderle et al., 2023). These habitat characteristics, usually related to the structure of the vegetation (e.g., tree cover, shrub cover, epiphytes, herb cover), determine the quantity and quality of available feeding resources for birds, which can directly affect avian functional diversity in coffee farms (Alvarez-Alvarez et al., 2022).
The importance of insectivorous birds as pest control agents is well documented (Nyffeler et al., 2018), especially for shade grown coffee (Chain-Guadarrama et al., 2019; Greenberg et al., 2000; Milligan et al., 2016; Philpott et al., 2009). Birds can significantly reduce infestations of the coffee berry borer beetle (Hypothenemus hampei), an important pest for coffee (Martínez-Salinas et al., 2016). The pest-control services provided by birds can vary with body mass, foraging strategies, and strata (Salgado-Negret and Paz, 2016). Tiny foliage gleaners like warblers, have been documented to occupy coffee farms with higher levels of tree cover where they prey on coffee berry borers (Campos et al., 2023). Other insectivorous gleaning and flycatching birds prefer increased vegetation structure and heterogeneity expressed as canopy, understory, and herb cover (Cubley et al., 2020; Melo et al., 2020).
Bird ecology studies typically suffer from two important methodological limitations. First, the inherent statistical and conceptual challenges of using diversity indexes, and their associated losses of information derived from confounding parameters of species abundance and species composition described by Barrantes and Sandoval (2009). Second, a large percentage (77%) of biodiversity studies estimating abundance, occupation, or richness fail to adequately consider the effect of imperfect detection of species within their analyses (Kellner and Swihart, 2014). The failure to address imperfect detectability can lead to incorrect diagnoses of trends in both abundance and species distribution (Lahoz-Monfort et al., 2014), and bias resulting management recommendations (Mackenzie, 2006). The application of occupancy models corrected for detectability (MacKenzie et al., 2002) can help address these problems by estimating the detection probability of species and more accurately relating occupancy estimations with specific habitat characteristics (Bailey and Adams, 2005).
Our study was conducted in a coffee landscape located in Oaxaca’s Sierra Mixe region in southern Mexico. Here, our objective was to evaluate the effects of habitat characteristics related to coffee management on the occupancy of different functional groups of insectivorous birds and their potential ecosystem service provisioning. Moreover, by evaluating the effects of habitat characteristics associated with coffee habitat management strategies on the occupancy of different functional groups of insectivorous birds, we provide coffee producers with recommendations on how to maximize their capacity to control pests in coffee-dominated landscapes, providing relevant contributions to productive systems in Latin America.
We hypothesized that specific habitat characteristics determine the occupancy probability of insectivorous bird species within the studied landscape. In particular, we expected that higher structural diversity of tree, epiphyte, shrub, and herb cover, and higher tree and shrub richness, increase the occupancy of both foliage gleaners and flycatcher insectivores (Cruz-Angón and Greenberg, 2005; Moradi et al., 2010). Conversely, habitat characteristics associated with reduced structural diversity (e.g., high coffee dominance; Rodrigues et al., 2018) should decrease occupancy by these groups. Occupancy of medium sized foliage gleaners was predicted to increase with higher tree cover (Waltert et al., 2005).
To our knowledge, this is the first study that models the variation of bird occupancy in relation to habitat characteristics from a functional groups approach. It is also one of the first occupancy analyses that consider imperfect detectability of birds of productive systems in Latin America. Finally, it provides practical management recommendations that are feasible for coffee producers to apply and could enhance the birds’ pest control service on coffee farms, benefiting coffee production and bird conservation.
MethodsStudy landscapeOur research was conducted in Santa Cruz Ocotal Mixe in the state of Oaxaca, Mexico (Fig. 1). Indigenous Mixe communities use the land mainly for coffee production, their principal cash crop. However, the study site is highly modified by human intervention, which has created a heterogeneous landscape with no clear differences among land uses that are interspersed across the territory; this condition promotes sudden habitat changes within short distances. The landscape, therefore, corresponds to a “coffee landscape” sensu Harvey et al. (2021), as it represents a complex mosaic of habitats where coffee is interspersed with other land uses such as secondary forest patches and various types of crops for self-consumption. Shade coffee itself is often diversified with different tree species and crops interspersed between coffee plants, and is the most important crop in this landscape due to its potential economic benefits for farmer communities and dominance in terms of overall area compared to other land uses. Land uses include coffee farms, urban settlements (land dedicated to housing where no coffee or other plantations occur), orchards (bananas, mango, tamarind, guava, avocado trees, or small sugar cane plots), secondary forests (pine-oak, evergreen, and cloud forest remnants), and milpas (plots dedicated to plant corn and beans). Coffee production is dominated by smallholders (0.55–2 ha). Approximately 70% of coffee farms are certified organic. Coffee is grown as a rustic or traditional polyculture shaded system (Moguel and Toledo, 1999) with a canopy dominated mostly by native trees at least 10 m tall and scattered fruit trees. The coffee harvest season extends from December to March, while pruning, weed removal, replanting, and shade regulation is performed at the end of the harvest season in April. Annual crops are produced from June through December across the region.
Habitat characteristics Location of the study area highlighted in Oaxaca (dark gray), southern Mexico (16°58")
To evaluate the effects of habitat characteristics on the occupancy of insectivorous birds, we randomly established 150 point-count stations with a 30 m fixed radius, separated by a minimum distance of 200 m along established trails in different land management contexts within the study area (i.e., coffee farms, urban settlement, orchards, secondary forests, milpas; Fig. 1) to ensure data independence. In each point-count we measured a total of seven habitat characteristics (i.e., tree cover, shrub cover, herb cover, coffee dominance, epiphytes, tree richness, shrub richness) and one local abiotic factor (elevation) for evaluations of occupancy. Their detailed descriptions are included in Table 1.
Description of the seven habitat characteristics and one local abiotic factor that were used for modelling the occupancy of insectivorous birds. Local abiotic factors are represented by elevation. Previously known effects of the habitat characteristics and abiotic factors on birds were noted with symbols. + = the effect is usually ecologically beneficial; − = the effect is usually ecologically detrimental; * = the effect is highly variable depending on the local context.
| Habitat characteristics/local abiotic factor | Description | Effect on birds |
|---|---|---|
| Tree cover | Visual estimation of the percentage of the point-count station covered by the canopy of trees taller than 5 m. | + sensuPhilpott and Bichier (2011) |
| Shrub cover | Visual estimation of the percentage of the point-count station covered by shrubs less than 5 m high. | + sensuOrtega-Álvarez et al. (2018) |
| Herb cover | Visual estimation of the percentage of the point-count station covered by herbaceous plants. | |
| Coffee dominance | Index from 0 to 1 resulting from the calculation of the total number of coffee plants divided by the number of all woody plants (shrubs, trees and coffee plants) in point-count stations. | - sensuRodrigues et al. (2018) |
| Epiphytes | Percentage of the surface of the trunk and large branches covered by epiphytic plants (e.g., vines, bromeliads, orchids, mistletoes) of ten trees within each point-count station, selecting two for each cardinal direction from the center to the outer limit of the station and two in the center of the station. Percentages were averaged to obtain a single value per station. | + sensuCruz-Angón and Greenberg (2005) |
| Tree richness | Number of tree species within the point-count station. | + sensuPhilpott and Bichier (2011) |
| Shrub richness | Number of shrub morphospecies within the point-count station. | + sensuDíaz (2006) |
| Elevation | Altitude of each point-count station that was measured with a GPS in meters above sea level. | * |
Habitat characteristics were selected considering their ecological relevance for regional birds (Ortega-Álvarez et al., 2018) and their importance in coffee farm management (i.e., coffee dominance; Rodrigues et al., 2018). All visual estimations of the habitat characteristics were performed by the same observer to avoid biases. Coffee dominance was particularly important for our study given that coffee plantations were dominant across the landscape, have important implications for the configuration of avian communities, and are directly associated with habitat management (Gregory and Strien, 2010; Philpott and Bichier, 2011; Rodrigues et al., 2018). All habitat characteristics were measured once per season on the first visit to each point-count station; their means and standard deviations are provided in Appendix A in online supplementary material.
Due to the landscape heterogeneity and fuzzy limits among land uses, bird’s occupancy analyses were performed by considering habitat characteristics as explanatory variables instead of land uses. This approach has been previously used by other studies to predict responses of population parameters to habitat gradients (Anderle et al., 2023; Rodrigues et al., 2018), as well as to avoid inconsistent land use classifications (Moguel and Toledo, 1999) and ecologically uninformative results. For our study site, this design further enabled us to: a) identify specific habitat characteristics correlated with bird occupancy, b) assess the effect of habitat characteristics variability on bird occurrence, c) predict occupancy responses to several habitat scenarios across the landscape, including those present in coffee cultivation areas, and d) elaborate more detailed habitat management recommendations.
Bird surveys and functional groupsTo sample both resident and winter migratory species across the landscape, two sampling seasons were conducted, one in summer (June and July 2021) and the other in winter (February 2022). Survey months were deliberately selected to avoid regional migratory movements peaks and keep with the closed population assumption associated with single-season occupancy modelling. Sampling was conducted over a 10 min period at each point-count station from dawn until bird activity decreased notably (typically five hours) (Bibby et al., 1998; Ralph et al., 1993). Passing and hovering birds as well as highly mobile species (e.g., raptors, swifts, swallows) were not recorded to avoid double counts when changing point-count stations. Each point-count station was visited twice per season by a group of three different observers trained in the visual and auditory identification of local bird species, which generated a total of 6 observation sessions per station by season (2 visits by 3 observers = 6 observation sessions) to model occupancy. In each visit, the time between each observation (n = 3; i.e., one per observer) in each point-count station varied from 10 min up to 3 days, depending on the proximity of the next station, as suggested by Ruiz-Gutiérrez et al. (2019). This approach enabled us to avoid violations of the occupancy models assumptions of a closed population (Mackenzie, 2006). To prevent biases, the sequence of visits to each point-count station was determined randomly. Single-season occupancy models relied on species detection (detected or not detected) not abundance, and therefore observation sessions made on the same day (n = 6, two per observer per point-count station) did not result in the overestimation of species abundance by double counting of birds, but allowed to establish detection histories required for occupancy estimations (Mackenzie, 2006).
Selected species were subsequently classified into different functional groups of insectivorous birds. We formally constructed such functional groups with a Similarity Profile Analysis using the average cluster method of Euclidean distance from the R package “clustsig” (Whitaker and Christman, 2010). To run this analysis, we considered several species traits defined by field observations and information from the Birds of the World data base (Billerman et al., 2022). Included traits were: main foraging strata (understory or canopy), main foraging strategy (foliage gleaner, bark gleaner, or flycatcher), and body size (tiny <15 g, small 16–35 g, medium 36–60 g, large 61–120 g, and extra-large >120 g). The details of traits classifying species are available in Appendix B.
Data analysisPrior to constructing the occupancy models, we performed Pearson correlation tests between habitat variables to avoid multicollinearity. When a pair of habitat characteristics were highly correlated (p values above 0.6 or below −0.6), one variable was eliminated keeping the variable which represented fewer challenges for habitat management practices (easier to measure and regulate).
Occupancy modelling was restricted to species from the bird community that could make greater contributions on arthropod pest control across the landscape (Sekercioglu, 2012). We only analyzed species that: a) exhibited a diet of at least 30% insects (Billerman et al., 2022), b) belonged to a taxonomical family previously associated with some type of insect pest control (Michel et al., 2020), and c) were abundant (at least 49 detections) during our surveys. Single-season occupancy models that consider the effect of detectability and habitat covariates on parameter estimation were run for each species using the “unmarked” package of R (Fiske and Chandler, 2011). In particular, we used the “occu” function, which fits the single season occupancy model of MacKenzie et al. (2002) based on zero inflated binomial models (MacKenzie, 2006); logit link was used to model covariates for both occupancy and probability of detection. For a detailed description of occupancy estimation, please see MacKenzie (2006) and MacKenzie et al. (2002). Models were run separately by season, which enabled us to comply with the closed population assumption of the statistical technique and account for seasonal variability in: a) habitat characteristics, b) pests phenology, and c) the functional composition of avian communities. Modeling was performed on species with a minimum of 49 detections in the field to ensure sufficient data for fitting reliable models. To optimize the fit of the models to available data, we estimated probability of detection as a nuisance parameter (Guillera-Arroita et al., 2010), since the performance of the detection method was not of interest for our study. In this way, habitat relationships focused on occupancy estimation while controlling for the effect of imperfect detectability in our models. Occupancy models were constructed for each species considering the additive (no interactions) effect of a maximum of three covariates per model to avoid overparameterization and facilitate the interpretation of results. The three covariates included in the models were chosen by considering all the possible combinations of the total pool of the eight non correlated habitat characteristics measured for this study. A null model where no effect of habitat characteristics is considered was fitted to subsecuently evaluate model fit with different combinations of habitat characteristics.
Model selection was performed using multiple models and an information-theoretic approach. We based model selection on the second order Akaike’s Information Criterion (AICc) because it provides good model selections by adjusting for small sample sizes (Burnham and Anderson, 2002). Finally, we modeled and plotted both positive and negative relations between habitat characteristics and the associated occurrence of each bird species in each functional group (Appendix C). We did not model the response of species richness (total number of species in a functional group) to habitat characteristics because such approach may mask the individual responses of species to habitat traits. To generate response curves of the occupancy probability of each bird species as function of habitat characteristics we used model-averaged estimations.
To synthesize results for different functional groups of insectivorous birds we simultaneously graphed all the curves of bird species from the same functional group responding to a particular habitat characteristic. A minimum of two species per group were required to plot responses to a particular habitat characteristic. Management recommendations were constructed using critical values for each habitat characteristic on our figures, representing the threshold value above which occupancy of most species increased notably. We visually defined critical values as the inflection points where slopes became steeper sensu Ortega-Álvarez et al. (2018), indicating the value in which occupancy probability increased for most species. For cases where there was no inflection point (straighter tendency lines), the average value in the middle of the x axis of the habitat characteristic was considered as the critical value (Ortega-Álvarez et al., 2018). For cases with contrasting positive and negative occupancy probability responses, the critical value was considered as the value of the habitat characteristic where the average of most species’ occupancy was enhanced, as this represents a value where most species still show considerable occurrence. Elevation was not included in management recommendations as it cannot be modified by producers.
ResultsHabitat characteristicsANOVA tests showed that there were no significant differences among land uses for most habitat characteristics, with three exceptions: tree cover, herb cover, and coffee dominance (Appendix D). Tree cover was highest in the secondary forest, followed by coffee farms and orchards in both seasons. Herb cover was highest within the milpa, mainly during winter. Apart from this trend there was minimal variation in the mean values of habitat characteristics across seasons.
Bird surveys and functional groupsWe recorded a total of 177 bird species (129 in summer, 145 in winter) belonging to 12 orders and 36 families. Occupancy modelling was performed for 44 insectivorous bird species (Appendix E), of which five were summer migrants, six were winter migrants, and 33 were full year residents for the area. All species were considered habitat generalists as detections of forest specialist species were scarce. The Similarity Profile Analysis identified six functional groups of insectivores at the cutoff of 46% of dissimilarity of Euclidean distance: large foliage gleaners, species between 60 g and 129 g foraging in both understory and canopy; tiny foliage gleaners, species smaller than 15 g foraging on both strata; small and medium foliage gleaners, species between 15 g and 60 g foraging on both strata; small canopy flycatchers, species between 15 g and 35 g foraging on the canopy; bark gleaners, species between 35 g and 120 g foraging in tree bark at both strata; and canopy flycatchers, species between 15 g and 35 g foraging in the canopy (except for Myiopagys viridicata weighing less than 15 g but feeding primarily in the canopy) (Fig. 2).
Occupancy analyses
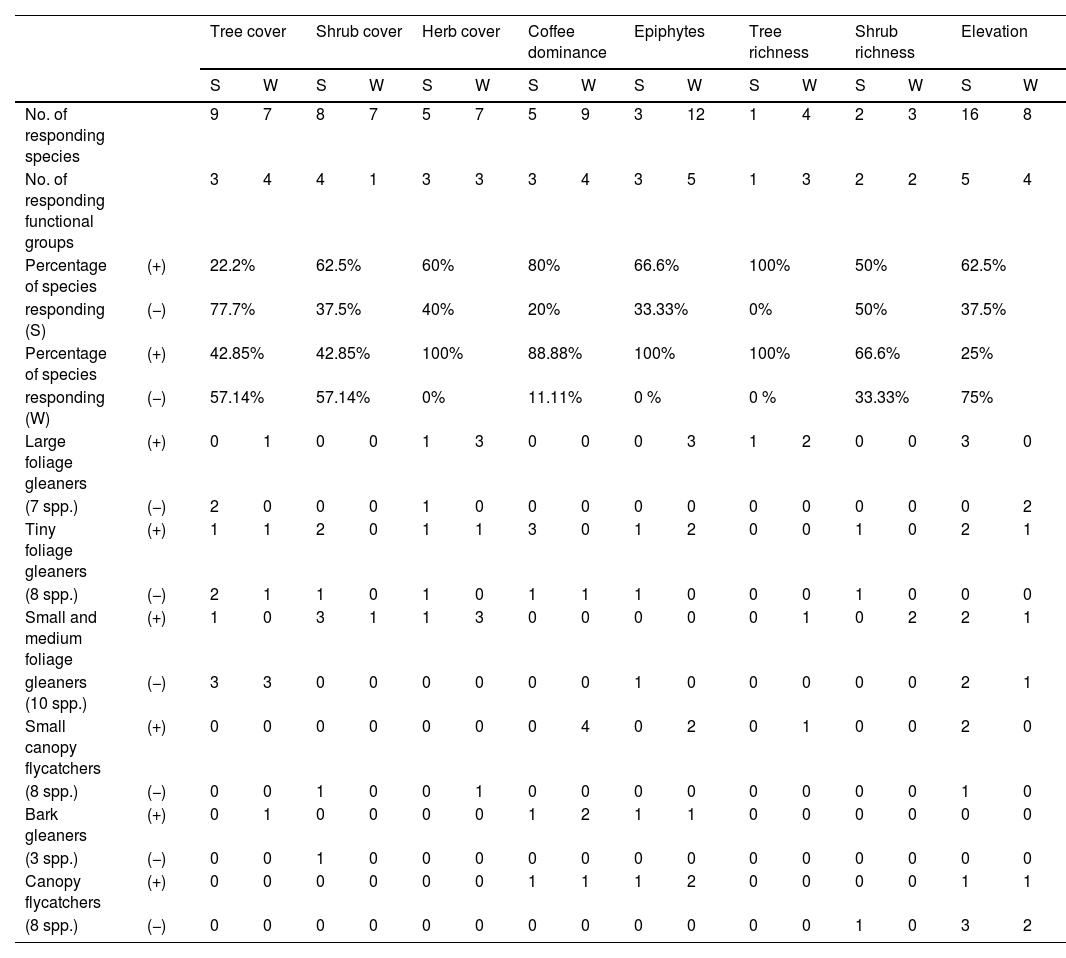
From the pool of 177 recorded species, we filtered for bird species with the restrictions noted in our data analysis section. After analysis, the occupancy of a total of 34 species in winter, and 22 in summer, was explained by habitat covariates (Appendix F). During summer, shrub cover was the habitat characteristic associated with the occurrence of the highest number of functional groups: tiny foliage gleaners, small and medium foliage gleaners, small canopy flycatchers, and bark gleaners (62.5% of species with a positive response and 37.5% of species with a negative response) (Table 2).
Number of species, number of functional groups, percentage of species, and number of species per functional group that responded to measured habitat characteristics in two seasons, summer (S) and winter (W). Positive (+) or negative (−) relationships are depicted.
| Tree cover | Shrub cover | Herb cover | Coffee dominance | Epiphytes | Tree richness | Shrub richness | Elevation | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S | W | S | W | S | W | S | W | S | W | S | W | S | W | S | W | ||
| No. of responding species | 9 | 7 | 8 | 7 | 5 | 7 | 5 | 9 | 3 | 12 | 1 | 4 | 2 | 3 | 16 | 8 | |
| No. of responding functional groups | 3 | 4 | 4 | 1 | 3 | 3 | 3 | 4 | 3 | 5 | 1 | 3 | 2 | 2 | 5 | 4 | |
| Percentage of species | (+) | 22.2% | 62.5% | 60% | 80% | 66.6% | 100% | 50% | 62.5% | ||||||||
| responding (S) | (−) | 77.7% | 37.5% | 40% | 20% | 33.33% | 0% | 50% | 37.5% | ||||||||
| Percentage of species | (+) | 42.85% | 42.85% | 100% | 88.88% | 100% | 100% | 66.6% | 25% | ||||||||
| responding (W) | (−) | 57.14% | 57.14% | 0% | 11.11% | 0 % | 0 % | 33.33% | 75% | ||||||||
| Large foliage gleaners | (+) | 0 | 1 | 0 | 0 | 1 | 3 | 0 | 0 | 0 | 3 | 1 | 2 | 0 | 0 | 3 | 0 |
| (7 spp.) | (−) | 2 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 |
| Tiny foliage gleaners | (+) | 1 | 1 | 2 | 0 | 1 | 1 | 3 | 0 | 1 | 2 | 0 | 0 | 1 | 0 | 2 | 1 |
| (8 spp.) | (−) | 2 | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| Small and medium foliage | (+) | 1 | 0 | 3 | 1 | 1 | 3 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 2 | 2 | 1 |
| gleaners (10 spp.) | (−) | 3 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 2 | 1 |
| Small canopy flycatchers | (+) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 2 | 0 | 1 | 0 | 0 | 2 | 0 |
| (8 spp.) | (−) | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Bark gleaners | (+) | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 2 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| (3 spp.) | (−) | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Canopy flycatchers | (+) | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 2 | 0 | 0 | 0 | 0 | 1 | 1 |
| (8 spp.) | (−) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 3 | 2 |
As shown in Fig. 3, occupancy of a particular number of species from each functional group was influenced by habitat characteristics. For instance, shrub cover was positively associated with all tiny, small, and medium foliage gleaner species, with the only exception being Troglodytes aedon, a tiny foliage gleaner adapted to rocky habitats with little to no vegetation. Tree cover was negatively associated with 7 out of 9 species from all three groups of foliage gleaners that responded to this trait, both species of large foliage gleaners, three small and medium foliage gleaners and two tiny foliage gleaners (Fig. 3). Herb cover was correlated both positively and negatively with the occupancy of large foliage gleaners (1 out of 2 species, respectively) and tiny foliage gleaners (1 out of 2 species). Coffee dominance was positively associated with 2 out of 3 tiny foliage gleaners, excluding Pheugopedius maculipectus.

Projected response of the occupancy probability of each insectivorous bird functional group to habitat characteristics during summer and winter. Each line represents a different insectivorous bird species. The line type, the color of the line, and the bird icon depicts the functional group of the species.
During the winter, tree cover negatively affected small and medium foliage gleaners (3 out of 3 affected species), and tiny foliage gleaners (1 out of 2 affected species) with the exception being Basileuterus culicivorus (Fig. 3). Herb cover was positively associated with small and medium (3 out of 3 affected species), and large foliage gleaners (3 out of 3 affected species). Coffee dominance had a positive relationship with three functional groups: bark gleaners (4 out of 4 affected species), small canopy flycatchers (4 out of 4 affected species), and tiny foliage gleaners (1 out of 2 affected species), with the exception of Basileuterus rufifrons. Epiphytes were positively correlated with the occurrence of every species affected by this trait from a total of five functional groups: tiny foliage gleaners (3 species), small canopy flycatchers (2 species), large foliage gleaners (3 species), canopy flycatchers (3 species), and bark gleaners (1 species) (Appendix E). Finally, tree richness positively affected the occupancy of large foliage gleaners (2 out of 2 species), whereas shrub richness was positively correlated with the occupancy of small and medium foliage gleaners (2 out of 2 affected species).
The estimated critical values where most functional groups increased their occupancy probability for each habitat characteristic are shown in Appendix A. Tree cover only exceeded its proposed critical value within coffee farms and secondary forests in both seasons. Shrub cover was higher than its respective critical value for both seasons in milpas and orchards, but only higher in coffee farms during summer. Herb cover only exceeded its proposed critical value within milpas during winter. Epiphytes never reached or exceeded their proposed critical values; highest values were found in orchards during summer and in secondary forest during winter.
DiscussionThe minimal variation of habitat characteristics values of land uses showed that directly assessing habitat characteristics as determinants of bird occupancy was a better approach to predicting species occupancy probabilities than land uses. We avoided modelling abundance/diversity of avian groups because information on individual species (e.g., detectability) might be lost during such processes. Instead, we modeled occupancy of each species by considering their particular probability of detection, which enabled us to differentiate between occupancy and detectability habitat relationships. Finally, with these results, we were able to propose habitat management recommendations for enhancing the occupancy of focal functional groups as a whole. We found more relationships of habitat characteristics influencing the occupancy probability of birds from functional groups in winter than in summer. These findings can be a result of the resources that the rustic coffee farms can provide, as birds may depend on them to survive during this season. Coffee is a perennial crop offering flowers and fruits year round, but more importantly, farms in our study site maintain forest-like characteristics such as high tree cover and a complex vertical structure that might provide important micro-habitats for both resident and wintering insectivore migrants (Bakermans et al., 2012). Greater positive influences of habitat characteristics on insectivore occupancy in winter could be related to changes in prey preferences of resident insectivorous birds between seasons (Jedlicka et al., 2021). This might also be explained by the higher number of migratory species that visit the coffee landscape during the winter.
Several studies have documented increased abundance of insectivore birds with increased canopy cover within coffee plantations and forests (Gordon et al., 2007; Philpott and Bichier, 2011). In contrast with these findings and our own hypothesis, our results showed that tree cover decreased occupancy of most small-medium (5 out of 6) and tiny foliage gleaners (2 out of 3) that responded significantly to such habitat characteristic during both seasons. Most of the large foliage gleaners that were associated with tree cover (2 out of 3 species) also responded negatively with this habitat characteristic. The negative relation of small gleaning birds with tree cover was also observed by Bakermans et al. (2012), and may be due to trees shading out lower vegetation layers, which can be detrimental to the bird groups that depend on insects tied to understory foliage (Bouvet et al., 2016). The reliance of tiny, small, and medium foliage gleaners on shrub cover and richness highlights the importance of maintaining shrubs biomass and diversity in managed ecosystems for foliage gleaning insectivores (Ortega-Álvarez et al., 2018), as shrubs can act as diverse food sources for prey species and provide additional perching, refuge, and nesting sites for birds (Díaz, 2006).
As we hypothesized, herb cover was a good predictor of occupancy by small, medium, and large foliage gleaners during winter. This result supports the findings of Leyequién et al. (2010) for similar species. Herb cover increases vertical vegetation structure, supplying habitat and food resources for insectivores that forage at lower feeding heights and enhances bird functional diversity in forest fragments (Melo et al., 2020). Herbaceous plants could also be functioning as arthropod refuges, thus being important food sources for birds foraging in our landscape, especially at times when vegetation in other strata or land uses may be reduced (e.g., milpas; Smith et al., 2015).
Although coffee dominance is often associated with increased management intensity, this does not necessarily affect insectivorous bird abundance (Rodrigues et al., 2018). Contrary to our predictions, we observed that several functional groups responded positively to this habitat characteristic. This result may be partially explained by three observations: 1) most organic coffee farms in the study region use a minimum amount of insecticides; 2) farms are embedded within secondary forest vegetation, which can increase bird occurrence (Galloway et al., 2021), and 3) there is a lack of primary forests in the area, converting shade grown coffee farms (with high percentages of tree cover; mean = 54%) into alternative habitat refuges for insectivores (i.e., bark gleaners; Arriaga-Weiss et al., 2008).
The probability of occupancy by tiny foliage gleaners also increased with coffee dominance. This is of particular interest for management, as members of this group have been identified as effective pest control agents (Karp et al., 2013), in particular species of the Parulidae family, that are consumers of coffee berry borers (Kellermann et al., 2008; Sherry et al., 2016). Even though the coffee berry borer can accumulate in large numbers inside growing and fallen berries during coffee berry production in winter (December–March; Baker and Barrera, 1993), our results are particularly relevant for managing the coffee berry borer at the end of its breeding season, which occurs in the beginning of summer (Baker and Barrera, 1993). The higher occupancy probabilities response from tiny, and small and medium foliage gleaners to both coffee dominance and shrub cover during the summer becomes more relevant to the pest control service against coffee berry borer, as females are only available for predation when they disperse to find more fruits during the beginning of the rainy season (summer) (Damon, 2000). In this sense, promoting shrub cover, especially during summer, may benefit local pest control services provided by tiny, small, and medium foliage gleaners as they represent the functional groups that are most effective in controlling insects in coffee plantations (Kellermann et al., 2008; Martínez-Salinas et al., 2016). We noted that this service could be complemented during the winter by the increased occupancy of tiny foliage gleaners, bark gleaners, and small canopy flycatchers with coffee dominance. Even though the coffee borer is not available for bird predation during this time, birds from these functional groups can also feed on different types of pest-insects for coffee plantations, such as the coffee leaf miner and plant hoppers (Borkhataria et al., 2006).
The occupancy probability of all canopy flycatchers, large foliage gleaners, and tiny foliage gleaners increased in areas with high levels of epiphytes during the winter. These results are consistent with evidence suggesting that epiphytes play a significant role in creating nesting and refuge resources for these birds while promoting the diversification of invertebrate refuges that enhance the availability of prey for insectivorous birds (Cruz-Angón and Greenberg, 2005), thus helping to increase potential pest control services within coffee farms.
Management recommendationsAn increased number and diversity of insectivorous birds can enhance pest control services in coffee farms, specially by foliage gleaning species from the Parulidae family and flycatching birds from the Tyrannidae family (Kellermann et al., 2008; Martínez-Salinas et al., 2016; Sherry et al., 2016). Given that occupancy should be positively correlated with abundance (MacKenzie, 2006), management recommendations based on the thresholds of specific habitat characteristics may be useful for simultaneously increasing insectivores’ occupancy and pest control service across the study landscape. Nevertheless, we recognize that pest control is a complex ecosystem service that can involve multi-trophic interactions between service providers and agricultural pests that might not always result in higher yields by enhancing habitat conditions for birds in the landscape (Díaz-Siefer et al., 2022). For example, depending on the study system, a trade-off between pest predation and pest predation alleviation due to birds consuming other non-pest, pollinating, or even other pest controlling invertebrates, could result in a dampened ecosystem service (Garcia et al., 2021). In contrast, for coffee agroecosystems, a complementary effect has been observed between bird pest control and pollination services, as both have shown to increase the quantity and quality of coffee fruits respectively without negatively affecting one another (Classen et al., 2014) and even creating positive synergies (Martínez-Salinas et al., 2022). Further research on this topic in the same region may help shed light on multi-trophic interactions and their effect on pest control service.
Given that coffee farming in Latin America is usually practiced in diverse and heterogeneous landscapes (Harvey et al., 2021), our findings suggest that the use of habitat characteristics can be a practical approach to determine avian-habitat relationships. As birds do not exclusively occupy coffee farms, it is important to measure these characteristics throughout the landscape to identify their associated values and possible levels of specific ecosystem service provisioning (e.g., pest control). In this sense, we identified specific critical values of habitat characteristics that might contribute to increase the occupancy probabilities of different insectivorous birds and ultimately enhance their potential pest control service. Specific management recommendations related with such critical values include maintaining a tree cover around 45% inside coffee farms by favoring tree pruning over removal, and planting more native tree species or allowing them to establish naturally. We realize that the relation between shade and coffee yields is important for producers, as it can vary between coffee cultivars (Koutouleas et al., 2022). However, we expect that our proposed level of 45% can ultimately be beneficial for coffee yield production, as this tree cover can favor pest control provision by increasing the occupancy probabilities of tiny, and small and medium foliage gleaning insectivores. Based on our results, we recommend promoting or propagating epiphytes’ cover above 27% on shade coffee trees, suggesting an important threshold versus previous studies by Cruz-Angón and Greenberg (2005) contrasting epiphytes presence-absence only. We also recommend preserving a native shrub cover of 48%, and herb cover of 50% within farms or along their borders, as such values may be particularly important in benefiting foliage gleaners and all flycatcher functional groups. Avoiding epiphyte removal, identifying the epiphyte and shrub species native to the area, and cultivating them in or around coffee farms could be important in this regard. Keeping a diverse tree and shrub richness of 13 and 17 species respectively might also be important for small, medium, and large foliage gleaning insectivores. The presence of these insectivores could be promoted by diversifying crops inside farms or establishing living fences. Although marked dominance by coffee may favor the occupancy of tiny foliage gleaners, small canopy flycatchers, and bark gleaners, dramatic increases should be avoided as they typically require reductions in the number or diversity of shade trees that are also important factors in insectivore abundance and are typically associated with an increased use of agrochemicals.
Our results regarding the medium levels of tree cover that increase the occupancy probability of most foliage gleaners, support the current levels of tree cover (e.g., 40%) required by most conservation-based certifications (Rainforest Alliance, Smithsonian Bird Friendly). However, shrub, herb, and epiphytes cover, as well as tree and shrub species richness should be considered when seeking to maximize ecosystem functionality. The absence of unmanaged forests in the landscape should also be a major concern. Logging in adjacent plots of forest may lead to a reduction of insectivorous birds abundance (Vergara et al., 2021) and to the loss of forest-dependent species, which in turn could ultimately increase insect herbivory in a trophic cascade (Peter et al., 2015).
ConclusionsThe sustainability of coffee production can be improved if management practices that enhance production also act synergistically with biodiversity conservation strategies. In this sense, coffee certifications such as Organic, Rainforest, Fair Trade, and Smithsonian Bird Friendly, might benefit from positive feedbacks in management strategies that generate increased earnings for growers by promoting practices conserving or restoring services dependent on vegetation structure such as pest control (Bray and Neilson, 2017). To help counter current threats to sustainable shaded coffee systems, our study not only identified habitat characteristics that promote biodiversity, but also offers ways to improve the balance between conservation and sustainable farms production by focusing on functional groups important to the provision of pest control services. Occupancy modeling represent a robust analytical tool to determine species-habitat relationships and that may help to achieve this balance. Additional studies should be performed to evaluate the role of uncommon species, monitor the effects of land use change over time, examine multi-trophic interactions, evaluate the effect of landscape traits on insectivores’ occupancy, and combine occupancy modelling with experimental evaluations on pest removal by birds.
We are thankful to the community and the Unidad Productiva of Santa Cruz Ocotal Mixe, for formally allowing this research to be implemented, for supporting and hosting our team during both field seasons, and for making this project possible. We thank Antonino Solano Francisco, Alejandro Pedro Francisco, Eustacia Pablo García, Eduardo Sainz, and Ana Tejero for their assistance in the field. We appreciate the artistic contributions of Paulina Hernández Rojas for improving the design of the figures of the paper. We acknowledge the comments and suggestions provided by five anonymous reviewers, who increased the quality of our work. We are grateful for the financial support provided by Neotropical Bird Club with a medium grant award and Idea Wild for the completion of our field work. RO-A gratefully acknowledges the support from the DGAPA-UNAM Postdoctoral Fellowship 2021-2022 during which this article was performed. This work was submitted as a fulfillment of JAF-A to obtain the MS degree in Posgrado en Ciencias of the Instituto de Ecología A.C. where JAF-A received a MS scholarship from CONAHCYT (1079265).
The following are Supplementary data to this article:

