





Human-caused habitat loss and fragmentation have significantly impacted the natural environments of large carnivores, altering their movement patterns and increasing risks such as hunting and road collisions. This study aims to understand the movement of jaguars (Panthera onca) through forests, agriculture of varying patch sizes, their distances to these structures, and roads and drainages. By analyzing movement speed, revisits, time spent inside these structures, and the timing of the last visit, data from 54 GPS-tagged jaguars in South America reveal a pronounced tendency to revisit the edges of these landscape variables. Additionally, jaguars showed a stronger affinity for natural areas, spending more time in large forest patches and reducing their speed in natural drainages. Areas with extensive agriculture had fewer revisits, and jaguars moved faster near roads. These results demonstrate the level of tolerance and the dangers this species faces in a landscape with anthropogenic aspects. This comprehensive assessment of movement patterns and landscape use provides valuable insights into how landscape structure influences habitat preference and mobility rates, which is crucial for future jaguar conservation and management strategies.

The natural habitats of terrestrial mammals are being lost due to the expansion of human activities (Crooks and Sanjayan, 2006), and the resulting habitat fragmentation and loss of connectivity threaten mammal populations worldwide (Crooks et al., 2011; Crooks and Sanjayan, 2006; Tucker et al., 2018; Woodroffe and Ginsberg, 1998). Large carnivores are particularly susceptible to habitat loss and fragmentation given their large body sizes and extensive home ranges required to find sufficient food and shelter (Crooks, 2002; Thompson et al., 2021), their low densities, and their slow growth rates (Stier et al., 2016; Woodroffe, 2000). Moreover, the carnivores have a much higher chance of becoming extinct regionally in fragmented habitats due to human encounters or persecution compared to other species groups (Woodroffe and Ginsberg, 1998; Woodroffe, 2000; Csermak et al., 2022). Thus, it is essential to identify how patch sizes affects the movement patterns of carnivores to understand the species' adaptability and provide improved evidence to manage and conserve large carnivores in human-dominated landscapes.
Movement studies of large carnivores indicate how vital habitat connectivity is (Kramer-Schadt et al., 2011), as these species cross great distances and have extensive home ranges (Thompson et al., 2021). In continuous habitats, large carnivores are able to find food and reproduce. In contrast, they are often forced to explore anthropic areas for food in landscapes with small and scattered habitat patches (Dobson et al., 2006; Mortelliti and Boitani, 2008; Péron et al., 2017), which often leads to carnivore-human conflict and consequently, persecution by humans (Woodroffe, 2000; Csermak et al., 2022).
The jaguar (Panthera onca) is one of the large carnivores in the Neotropics that faces challenges due to habitat fragmentation and conversion of natural lands to agriculture, which also contribute to the reduction of its current distribution (de la Torre et al., 2018; Cullen et al., 2016; Haag et al., 2010; Zimmermann et al., 2021). Although jaguars prefer natural vegetation areas (Alvarenga et al., 2021; Conde et al., 2010; Morato et al., 2018a), they also use agriculture, peri-urban areas and roads (Alegre et al., 2023; Colchero et al., 2011; Cerqueira et al., 2021; Morato et al., 2018a) which increase their risk of mortality from road collisions and encounters with humans (Alegre et al., 2023; Cerqueira et al., 2021; Cullen et al., 2016). Additionally, jaguars' natural tendency to stay near forest edges (Dos Santos et al., 2022; Alegre et al., 2024) may lead to interactions with human activities, particularly in modified landscapes (Zimmermann et al., 2021). However, quantitative studies exploring jaguar movement patterns in landscapes modified by agriculture are scarce. By modeling jaguar movement responses to habitat modification, we can identify the landscape variables that impact the species and determine the degree of landscape modification they can tolerate (Bastille‐Rousseau and Wittemyer, 2021; Kays et al., 2015). This knowledge is essential for developing effective conservation strategies and management measures that promote coexistence between jaguars and human activities and ensure the long-term survival of this iconic species.
Here, we deepen the knowledge about the movement behavior in human-dominated landscapes where the jaguar lives, a top predator in Central and South America with a near-threatened category by the IUCN (Quigley et al., 2017). Our primary aim is to determine the movement patterns of jaguars concerning the structure of forest patches and agriculture areas, as well as the distances within and outside these structures. Additionally, we analyze how the proximity to drainage and roads influences these movement patterns. We considered movement patterns such as revisit (number of visits), time inside (time that the jaguar spent in one of these landscape structures), time since the last visit (of the structures), and speed as response variables within our models, translating the frequency of use of areas into more detailed return metrics. In addition, we determined at what depth jaguars enter agriculture and forest areas and how long they remain within them. We expected that, as a carnivore that depends on forested spaces for its survival (De Angelo et al., 2013, 2011), the jaguar tends to spend more time inside the forest and avoids spending long time in agriculture or near roads. Revisits are frequently carried out in small patches of forests and agriculture, used in the movement of the jaguar and search for resources and expansion of its home range (Conde et al., 2010; Thompson et al., 2021). Finally, we expected the jaguar movement pattern to respond negatively to anthropic structures (such as agriculture and roads).
Given the exploratory nature of this study and the limited peer-reviewed literature, we did not provide specific expectations for every variable combination. Instead, our approach was to generate a comprehensive understanding of jaguar movement patterns, which can inform more specific hypotheses in future research. If there are no specific expectations for some relationships, we aimed to present results regarding movement patterns relative to natural and anthropogenic variables, which provides clearer and more concise insights.
MethodsJaguar data and movement patternsWe selected 54 individuals from the Jaguar GPS open database (Morato et al., 2018b) distributed in South America (Fig. 1). The criteria to select individuals were: (1) regularity of data collection, meaning that during the specific sampling period, the individual's GPS data adhered to the established schedules, and (2) whether the fixed rate was 4 h or it could be resampled to 4 h (see Table A in Supplementary material). To keep all individuals standardized across different regions, we resampled data to 4-h intervals and analyzed the speed of each individual with ctmm package (Fleming and Calabrese, 2022). We also calculated revisitation metrics with the recurse package (Bracis et al., 2018), establishing a circle of 250 meters radius as a neighborhood for analysis. The 250-meter radius represents the mean population distance traveled in 4 h (see Table A in Supplementary material). The number of trajectory segments entering and leaving the circle centered in each location determines the number of revisits (Fig. 2, 1B). Also, this formulation allows us to compute the time inside on each visit and the time elapsed since the last visit to a given neighborhood (Fig. 2, 1C, and 1D). All analyses were performed within the R environment (R Core Team, 2022).
 of all individuals in each region.")
Distribution of jaguar data used in this study: the numbers indicate the regions in South America and the barplot, shows the percentage of forest, agricultural, and other land cover in each region. The polygons represent the overlap of home ranges (calculated with Autocorrelated Kernel Density Estimation -AKDE) of all individuals in each region.
 Movement patterns used as response variables: (A) We measured speed from one point to another, and (B) Revisit is the number of trajectories made within a radius of 250 meters, (C) The time spent on each visit, considering entry and exit time, and (D) the time spent returning to the 250-meter radius (figure adapted from Bracis et al., 2018). (2) Environmental explanatory variables: land cover data was reclassified into binary layers, and the distance from drainage and from roads was calculated from vector shapefiles transformed into raster layers. The landscape metrics (Patch size and distance of the variables) were derived from the binary maps (forest/non-forest and agricultural and non-agricultural areas).")
Variables used in this study. (1) Movement patterns used as response variables: (A) We measured speed from one point to another, and (B) Revisit is the number of trajectories made within a radius of 250 meters, (C) The time spent on each visit, considering entry and exit time, and (D) the time spent returning to the 250-meter radius (figure adapted from Bracis et al., 2018). (2) Environmental explanatory variables: land cover data was reclassified into binary layers, and the distance from drainage and from roads was calculated from vector shapefiles transformed into raster layers. The landscape metrics (Patch size and distance of the variables) were derived from the binary maps (forest/non-forest and agricultural and non-agricultural areas).
We used 95% confidence intervals to assist in comparing descriptive data and movement patterns in each landscape structure, such as forest and agriculture. To calculate the confidence intervals for the means of each variable, we used the standard error, which quantifies the variability of the sample mean. We used the criterion of overlapping confidence intervals to determine whether the differences between the means were significant. Specifically, if the 95% confidence intervals of the means for a given variable in the two areas did not overlap, we considered the difference statistically significant. Conversely, if the confidence intervals overlapped, we interpreted the difference to be not statistically significant (Moore et al., 2009).
Environmental data and landscape metricsOur primary aim was to determine the movement patterns of jaguars across various landscape structures, including forests, agricultural areas, drainage systems, and roads, elements that significantly influence jaguar behavior (Alegre et al., 2023). In this study, we analyzed movement patterns considering the configurations of each landscape structure, using metrics such as patch size and distance, to allow for an easier understanding of how these landscape structures affect jaguar movement.
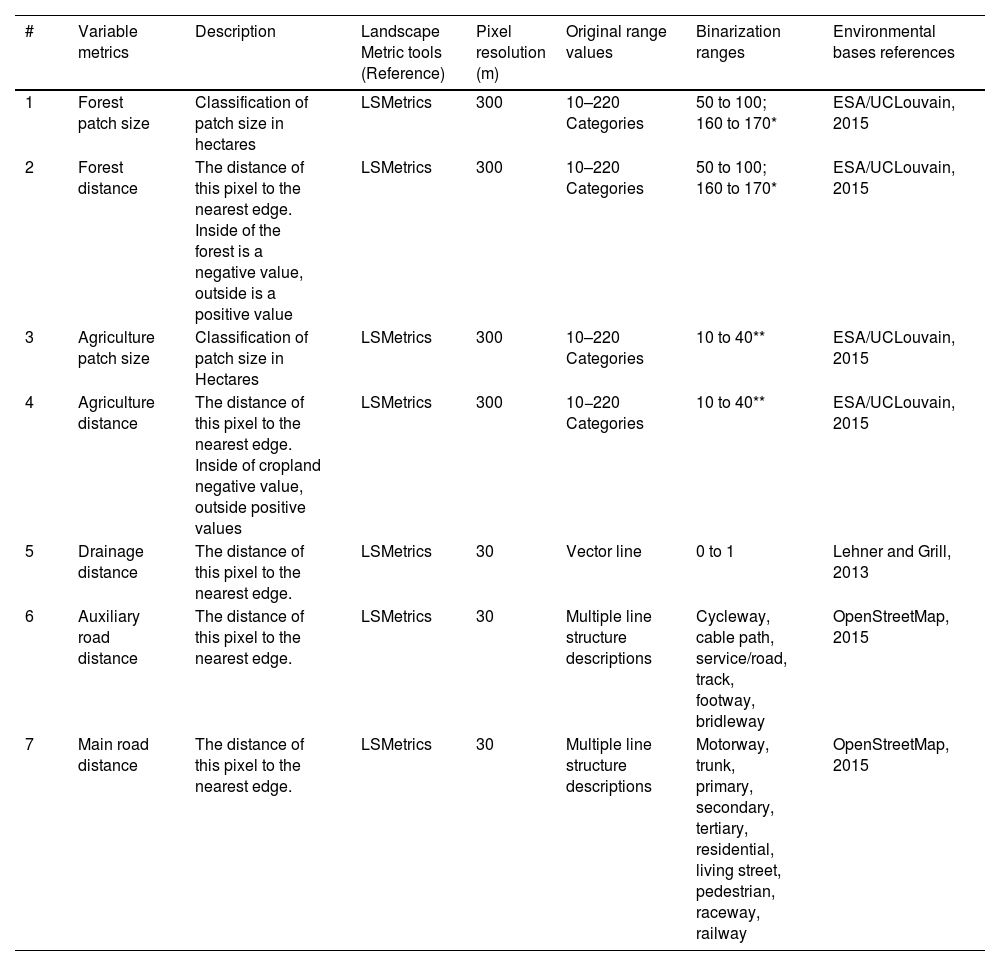
Therefore, we selected four environmental variables to calculate seven landscape metrics (Table 1). The raster for landscape structures, such as forests and agricultural areas, were obtained through the European Space Agency platform (ESA/UCLOVAIN, 2015). The vectors for roads (OpenStreetMap, 2015) and drainages (Lehner and Grill, 2013) were transformed into raster using QGIS 3.10.7 (QGIS Development Team, 2020) for subsequent analysis (Fig. 2). All rasters were reclassified in two categories (hereafter binarized) in the LandScape Metrics software (e.g., 1 = forest; 0 = non-forest; LSMetrics, Niebuhr, 2018). The ranges for this binarization for each environmental variable are detailed in Table 1, and its references for further exploration. For the binary variables of forest and agriculture, we used LandScape Metrics to obtain our metrics of patch size in hectares and distances both inside and outside of the patches. Specifically, the 'distance inside' represents the distance from any point within the patch (e.g., forest or agriculture) to the nearest edge of the patch, while the 'distance outside' refers to the distance from points outside the patch to the nearest patch edge. We opted to perform only distance outside metrics for the road and drainage variables, being linear structures.
The variable metrics information considered in this study includes a description of these metrics, the metrics tools used, the original pixel resolution, and the value ranges of our environmental databases from which these metrics were extracted. Also, the information includes the ranges used for binarization and, finally, the reference to the original environmental bases. The distance from main roads was removed from the final model because it exhibited collinearity with the distance to auxiliary roads, and also scarce in our study regions.
| # | Variable metrics | Description | Landscape Metric tools (Reference) | Pixel resolution (m) | Original range values | Binarization ranges | Environmental bases references |
|---|---|---|---|---|---|---|---|
| 1 | Forest patch size | Classification of patch size in hectares | LSMetrics | 300 | 10–220 Categories | 50 to 100; 160 to 170* | ESA/UCLouvain, 2015 |
| 2 | Forest distance | The distance of this pixel to the nearest edge. Inside of the forest is a negative value, outside is a positive value | LSMetrics | 300 | 10–220 Categories | 50 to 100; 160 to 170* | ESA/UCLouvain, 2015 |
| 3 | Agriculture patch size | Classification of patch size in Hectares | LSMetrics | 300 | 10–220 Categories | 10 to 40** | ESA/UCLouvain, 2015 |
| 4 | Agriculture distance | The distance of this pixel to the nearest edge. Inside of cropland negative value, outside positive values | LSMetrics | 300 | 10−220 Categories | 10 to 40** | ESA/UCLouvain, 2015 |
| 5 | Drainage distance | The distance of this pixel to the nearest edge. | LSMetrics | 30 | Vector line | 0 to 1 | Lehner and Grill, 2013 |
| 6 | Auxiliary road distance | The distance of this pixel to the nearest edge. | LSMetrics | 30 | Multiple line structure descriptions | Cycleway, cable path, service/road, track, footway, bridleway | OpenStreetMap, 2015 |
| 7 | Main road distance | The distance of this pixel to the nearest edge. | LSMetrics | 30 | Multiple line structure descriptions | Motorway, trunk, primary, secondary, tertiary, residential, living street, pedestrian, raceway, railway | OpenStreetMap, 2015 |
50-Tree cover, broadleaved, evergreen, closed to open (>15%); 60-Tree cover, broadleaved, deciduous, closed to open (>15%); 70- Tree cover, needleleaved, evergreen, closed to open (>15%); 80- Tree cover, needleleaved, deciduous, closed to open (>15%); 90-Tree cover, mixed leaf type (broadleaved and needleleaved); 100-Mosaic tree and shrub (>50%) / herbaceous cover (<50%);160 -Tree cover, flooded, fresh or brakish water; 170-Tree cover, flooded, saline water.
To avoid collinearity, we analyzed the correlation and variance inflation factor (VIF) between the environmental variables generated with the vifcor function from the usdm package (Naimi, 2015). We removed variables with a correlation > 0.7 and VIF > 3 (Table 1; Zuur et al., 2009). In the collinearity analysis, main roads were removed due to a correlation > 0.7 with auxiliary roads; we decided to remove this variable because it is also absent in one study area.
Models and diagnosticsWe fitted four generalized linear mixed models, one for each response variable; revisit, time spent, time since the last visit, and speed. We used agriculture patch size, agriculture distance, forest patch size, forest distance, drainage distance, and auxiliary road distance as explanatory variables for each model. We used individuals and regions as random variables of the intercept. All our models were fitted with the glmmTMB package (Brooks et al., 2017) in R (R Core Team, 2022). We used the t-family with an "identity" link function for the four models because it accommodates data with extreme values and provides robust estimates under these conditions (Gelman and Hill, 2007). The package glmmTMB` offers various extensions, including the ability to handle zero-inflated data, use a variety of response distributions, specify complex variance and correlation structures, address overdispersion, and fit models with censoring and truncation (Brooks et al., 2017).
To ensure the robustness and validity of our generalized linear mixed models, we performed several diagnostic tests using the DHARMa package in R (Hartig, 2022). These diagnostics included the Kolmogorov-Smirnov (KS) test, which evaluates the goodness of fit of the residuals by comparing the observed and expected distributions. A non-significant KS test indicates that the residuals follow the expected distribution, suggesting a good fit. Additionally, it includes the dispersion test to check for overdispersion in the models, which may indicate that the variance of the data is greater than predicted by the model. It also includes outlier testing to identify data points that deviate significantly from the model predictions. All models met the statistical assumptions for analysis of generalized linear mixed models, as confirmed by these diagnostic tests (See Appendix A in Supplementary Material).
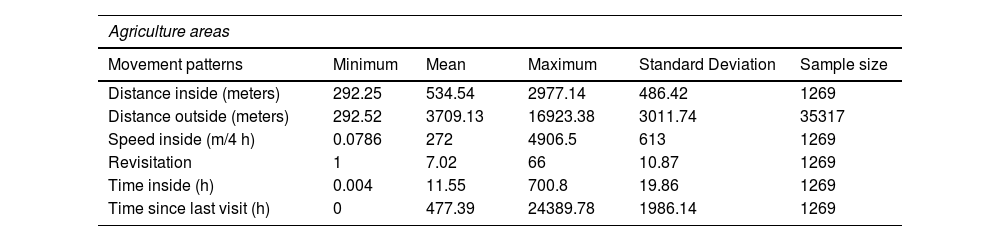
ResultsOur descriptive results indicated that the mean distance traveled by the jaguar inside agriculture was 534.5 meters, while in forest, it was higher at 986.5 meters. However, the mean distance the jaguar traveled outside agriculture was 3709.1 meters, whereas it was lower - 888.3 meters - outside forests. In contrast, some variables showed similar means; the speed in agriculture was 272 (m/4 h), whereas, in forest, it was slightly lower at 251(m/4 h). The revisitation rate was also similar, 7.02 in agriculture and 6.71 in forest. In terms of time inside agriculture, the mean was 11.6 h, while in forest, it was higher at 14.2 h. However, the mean time since the last visit in agriculture was 477.4 h, whereas in forest, it was 442.3 h (Table 2 and Figure A in Supplementary Material).
- -
Forest
Descriptive data of movement patterns of agricultural and forest areas.
| Agriculture areas | |||||
|---|---|---|---|---|---|
| Movement patterns | Minimum | Mean | Maximum | Standard Deviation | Sample size |
| Distance inside (meters) | 292.25 | 534.54 | 2977.14 | 486.42 | 1269 |
| Distance outside (meters) | 292.52 | 3709.13 | 16923.38 | 3011.74 | 35317 |
| Speed inside (m/4 h) | 0.0786 | 272 | 4906.5 | 613 | 1269 |
| Revisitation | 1 | 7.02 | 66 | 10.87 | 1269 |
| Time inside (h) | 0.004 | 11.55 | 700.8 | 19.86 | 1269 |
| Time since last visit (h) | 0 | 477.39 | 24389.78 | 1986.14 | 1269 |
| Forest | |||||
|---|---|---|---|---|---|
| Movement patterns | Minimum | Mean | Maximum | Standard Deviation | Sample size |
| Distance inside (meters) | 292.09 | 986.53 | 14987.8 | 1259.1 | 21468 |
| Distance outside (meters) | 292.09 | 888.32 | 9544.68 | 1367.45 | 15118 |
| Speed inside (m/4 h) | 0.057 | 251 | 9277 | 576 | 21468 |
| Revisitation | 1 | 6.71 | 67 | 8.56 | 21468 |
| Time inside (h) | 0.0002 | 14.22 | 1736.62 | 22.77 | 21468 |
| Time since last visit (h) | 0 | 442.3369 | 25074.88 | 1709.041 | 21468 |
In our models, we observed that the size of the forest patch does not have a significant effect on the movement speed of jaguars (β = 0.016, p > 0.05). In contrast, we observed that increasing the size of forest patches significantly reduces the number of revisits (β = −0.076, p < 0.01), which may be due to larger forest patches providing more extensive resources, such as prey and shelter, reducing the need for frequent returns. Additionally, the time jaguars spend within these larger forest patches increases significantly (β = 0.166, p < 0.001; Figure A in Supplementary Material), suggesting that larger patches offer a more suitable or rewarding environment, encouraging jaguars to remain within them longer. We registered that increasing the size of forest patches significantly reduces the time since the last visit (β = −0.527, p < 0.001). This implies that larger patches are likely revisited sooner, possibly because they serve as key habitats that jaguars rely on regularly for foraging, resting, or other essential activities.
Distance from the forest does not significantly affect jaguar speed (β = −0.065, p > 0.05). However, increasing the distance from the forest significantly increases the number of revisits (β = 0.057, p < 0.01); in fact, we observed that all the revisits are taking place at the forest edges (Fig. 3). Distance from the forest does not significantly affect the time inside that the jaguars spend in these areas (β = −0.01, p > 0.05). However, increasing distance from forests significantly affects increasing time since the last visit (β = 0.232, p < 0.05; Fig. 4, Table B in Supplementary Material).
- -
Agriculture

The number of visits across different distances of four land cover variables. Negative values indicate the distance within the variable, 0 is the edge, and positive values indicate the distance outside the variables. We provide figures for these variables separately in Figure C of the supplementary material.
 revisitation metrics (revisits, time inside, and time since last visit) and speed, with the different environmental variables (Values in Table B—Supplementary material).")
In our models, we observed that the size of the agriculture patch tends to reduce the movement speed of jaguars, although this effect was not statistically significant (β = −0.062, p ≈ 0.08). However, larger agriculture patches significantly reduce the number of revisits (β = −0.096, p < 0.001). Additionally, we found that larger agriculture patches significantly reduce the time inside that jaguars spend in these areas (β = −0.303, p < 0.001; Figure B in Supplementary Material). This result suggests that jaguars tend to avoid areas with larger agriculture patches. Furthermore, increasing the size of the agriculture patches significantly increases the time since the last visit (β = 0.316, p < 0.001), indicating that jaguars are not likely to return to these areas sooner.
Regarding distance to agriculture, we found that this distance does not significantly affect the movement speed of jaguars (β = 0.027, p > 0.05). However, increasing the distance from agriculture significantly reduces the number of revisits (β = −0.049, p < 0.05), with the jaguar's most frequent revisits occurring at the edge of agricultural areas (Fig. 3). Increasing the distance from agriculture significantly reduces the time inside of these areas (β = −0.47, p < 0.001). Finally, we found that increasing the distance from the agriculture significantly decreases the time since the last visit (β = −0.15, p < 0.05), indicating that jaguars are more likely to return sooner to areas further away from agriculture (Fig. 4, Table B in Supplementary Material).
- -
Drainage distance
In our models, we observed that increasing the distance to the drainage significantly positively affects the movement speed of jaguars (β = 0.246, p < 0.001). This result indicates that jaguars tend to move slowly near drainage areas. Furthermore, increasing the distance to the drainage significantly reduces the number of revisits (β = −0.27, p < 0.001). Still, like the other structures, we observed that revisits are evident at the edge of this structure (Fig. 3). This suggests that jaguars are less likely to revisit areas further away from the drainage.
Additionally, increasing distance to drainage significantly increased the time jaguars spent within these areas (β = 0.08, p < 0.001). This counterintuitive result could indicate that when jaguars venture further from drainage areas, they tend to stay longer. Finally, increasing the distance to drainage significantly reduces the time since the last visit (β = −0.91, p < 0.001), suggesting that jaguars return sooner to areas that are farther away from drainage (Fig. 4, Table B in Supplementary Material).
- -
Road distance
Regarding the roads, we found that increasing the distance to the roads significantly negatively affects the movement speed of the jaguars (β = −0.465, p < 0.01). This suggests that jaguars move faster near roads. Furthermore, increasing the distance to roads significantly reduces the number of revisits (β = −0.202, p < 0.05), indicating that jaguars revisit the roadside frequently (Fig. 3).
Furthermore, increasing the distance to roads significantly reduces the time inside that jaguars spend in these areas (β = −2.519, p < 0.001). This suggests that jaguars do not prefer to spend time in areas away from roads. However, distance to roads does not significantly affect time since the last visit (β = −0.441, p > 0.05; Fig. 4, Table B in Supplementary Material).
DiscussionOur results reveal that jaguars frequently visit the interface between forests, agricultural areas, roads, and drainages. The time between revisits and how long they stay inside these structures varies according to the size of the agriculture and forest patches, offering new insights into jaguar behavior in fragmented habitats. Furthermore, distance to these structures, such as forests, agriculture, drainage areas, and roads, significantly and variably affects jaguar behavior. Jaguar movement speed is differentially influenced by proximity to various landscape features: jaguars tend to move faster near roads and slower near drainage areas. At the same time, distance to forests and agricultural lands seems to have a less pronounced effect. These results contribute to a better understanding of what structural aspects within the habitat should be considered to manage natural habitats throughout the species distribution.
Jaguars are considered a crucial species for conserving natural habitats because they are closely associated with forested areas (Cullen Junior et al., 2013; Thornton et al., 2016; Morato et al., 2018a). Large carnivores require extensive areas with abundant natural prey (Cavalcanti and Gese, 2009; de la Torre et al., 2017; Thompson et al., 2021). Our results showed that the jaguar makes many revisits in small patches of forest and at the edge of the forest, which could suggest that it uses these areas for sustenance. This behavior might be driven by the variability in resource quality, where jaguars exploit lower-quality prey or easier hunting opportunities at these patches (Norris et al., 2010), as rodents (e.g., Agouti paca) are abundant in smaller forest patches (Norris et al., 2008). These smaller forest patches, also named “resource islands”, are created by trees and shrubs, which support higher biodiversity and greater ecological interactions compared to the surrounding grassland (Vetaas, 1992). Other larger prey, such as ungulates, frequently visit forest edges, influenced by landscape configuration (Lima et al., 2019; Norris et al., 2008; Ries et al., 2004), which can attract constant revisits by the jaguar. However, we also recorded that jaguars spend more time in larger forest patches than smaller ones, indicating that forests remain a safe area. This preference for larger patches likely reflects a strategy to optimize access to high-quality resources and avoid the risks associated with more anthropized areas (Cavalcanti and Gese, 2009; Conde et al., 2010; Morato et al., 2018a). Similar behavior is observed in pumas, which spend more time in larger patches and less in concentrated prey areas (Laundré, 2010). These findings support our expectation that jaguars will spend more time inside the forest, underscoring the critical importance of conserving large, contiguous forest patches. These areas not only serve as safe havens but are also essential for maintaining the ecological dynamics of jaguar populations, providing the necessary resources that are less available in anthropized environments.
We expected that jaguars would avoid agriculture; our results indicated a reduction in visits to larger agricultural areas and that it does not spend much time in them, which is an avoidant behavior in the species in proportion to the size of the anthropic structure. However, we recorded that most of the revisits were carried out on the edge of this structure, which might suggest that these edges provide easier access to opportunistic prey (Alegre et al., 2024; Esparza-Carlos et al., 2018) that may feed on crops or waste (Fitzgibbon, 1997; Dijak and Thompson, 2000; Bateman and Fleming, 2012; Ditmer et al., 2021). For example, the white-lipped peccaries, a vital prey of the jaguar, can use agriculture areas to feed, and due to this, conflict with humans have been registered in specific areas of Brazil (e.g. near the Emas National Park) where land use was mostly coverted for corn plantations (Lima et al., 2019; Csermak et al., 2022). This highlights how human structures and landscape modifications directly and indirectly alter predator-prey interactions (Patten et al., 2019; Berger, 2007; Murphy et al., 2021), sometimes leading to "empty forests" (Pires and Galetti, 2022), causing jaguars to leave the forest for search of resources.
Jaguars also frequently revisit natural drainage edges, which provide hydration and potential prey like capybaras and caimans (Azevedo and Murray, 2007; Azevedo and Verdade, 2012) in regions such as Pantanal wetlands. Furthermore, we found that they move slowly near the drainage, but the time spent near them is less, which delays the next visit. The drainage areas in many jaguar habitats are exploited and intervened by man (Ikeda-Castrillon et al., 2022), which could cause this transitory use.
Our results also indicated that jaguars frequently revisit the edges of the roads. Recent studies show that roads did not directly affect jaguar space use (Cerqueira et al., 2021). However, adverse effects of roads on jaguars' space use occurred indirectly in the same study because paved roads were associated with a low proportion of forest. Jaguars may also use the road positively as a resource for displacement (Alegre et al., 2023). Similar studies with large carnivores demonstrated road use according to traffic intensity (Kautz et al., 2022). However, roads, especially paved ones, pose significant risks due to high probability of wildlife collisions, leading to high mortality (Espinosa et al., 2018). These results underscore the importance of considering landscape structure in conservation strategies and highlight the complex interactions between jaguars and their fragmented habitats.
On the other hand, our study has some limitations that must be considered. One limitation is the heterogeneity in GPS data collection methods for different jaguars, which can introduce variability in the precision and resolution of the movement data. Additionally, our analysis did not account for the sex of the jaguars because our data did not include a uniform distribution of males and females across different environmental types. Only males were present in some areas, while in others, only females or a mix of both, making it challenging to analyze sex-specific differences in movement patterns. Another limitation is the reliance on auxiliary road data from OpenStreetMap (OSM), as we were unable to verify the accuracy of all mapped roads due to resource constraints. Future studies should consider validating these datasets, as this could enhance the precision of movement analysis. Finally, future research should develop movement patterns that differentiate between sexes, as this could provide more detailed information on how these landscape structures affect the jaguar population. Addressing these limitations in future research will enhance our understanding of the jaguar movement and contribute to more effective conservation strategies.
Conservation implications and conclusionsThe jaguar is a threatened species throughout its range, mainly in areas where it shares its habitat with humans (Castaño-Uribe et al., 2016). Our results could be a key to explaining one of the consequences determining the conflict with humans when using the same areas. Our results demonstrate that the jaguar frequently revisits the edges of forests, agriculture, roads, and drainages. It shows that the landscape's configuration and composition influence the jaguars' movement. Therefore, landscape structure and distance from anthropogenic disturbances affect habitat use. Large forest areas remain safe for the species despite their presence in anthropogenic structures. Studies show that, in addition to landscape structure, patch size plays a significant role, as larger patches can provide refuge and more resources. Moreover, the abundance of food resources, both within and outside these patches, is a key factor that could determine carnivore movements in fragmented landscapes (Mortelliti and Boitani, 2008).
To make informed decisions about species management and conservation, we must focus on buffer zones in natural habitats. Buffer zones are transitional areas between natural vegetation and human-modified lands, where land use is typically managed to reduce human impact on wildlife (Naughton-Treves et al., 2005). In many cases, these zones are subject to land use restrictions that can be legally enforced to protect wildlife and natural resources (Naughton-Treves et al., 2005; Paolino et al., 2016), and the way and timing of human use can alter the ecological interaction dynamics of large carnivores (Oriol‐Cotterill et al., 2015). Encouraging the implementation of buffer zones often requires a combination of policy measures, community engagement, and incentives for landowners to adopt conservation practices (Dudley, 2008). It is also crucial to consider the composition of shrubs and vegetation within these areas to provide optimal conditions for feeding and sheltering the prey of large carnivores, preventing them from seeking refuge near anthropogenic areas. These areas help maintain the dynamics between jaguars and their prey, ensuring that prey has sufficient cover and food resources to avoid approaching human zones, reducing the risk of conflicts between jaguars and humans. The protection and management of these areas can be improved through continuous monitoring and research to evaluate the effectiveness of conservation strategies and adjust policies as necessary.
Additionally, as our results reveal, buffer zones and "resource islands" that may represent an abundance of prey for the jaguar are crucial for the survival of this species. These regions, such as certain areas of the Amazon and the Pantanal in Brazil, provide a habitat rich in biodiversity where jaguar prey, such as agouti, peccaries, and capybaras, are abundant (Norris et al., 2008; Paolino et al., 2016). To effectively manage and conserve these resource islands, it is essential to implement strategies that include creating and maintaining biological corridors connecting different habitats Such corridors are likely to enhance the safe mobility of jaguars and improve their access to prey, while potentially reducing human-jaguar interactions and, consequently, conflict (Zeller et al., 2013). These strategies should also focus on protecting the vegetation and natural resources within the islands to ensure that jaguar prey has sufficient food and shelter.
Prey dispersal and the intensity of human disturbance in different regions may also influence jaguar movement within different contexts of landscape structure, making it difficult to decouple different effects. However, monitoring the dispersal strategies of diverse landscape-dependent populations could help understand how these populations respond to different landscape structures. Future studies should focus on how such revisit patterns will change, as landscape changes are dynamic and deforestation still occurs in many biomes across the Neotropics. Additionally, consider the insertion or maintenance of species that can maintain the ecological chain within them. A better understanding of landscape effects on predator-prey dynamics can provide additional insights into the components that drive these occupational strategies within various habitat structures.
This study and VBA received support from the São Paulo Research Foundation - FAPESP (processes number 2018/13037-3. 2020/07586-4). The co-authors received support from the São Paulo Research Foundation - FAPESP (processes number 2016/11595-3. 2019/04851-1. 2021/02132-8). JJT was partially supported by the program PRONII of the Consejo Nacional de Ciencia y Tecnología (CONACYT) of Paraguay. MCR thanks to the Sao Paulo Research Foundation - FAPESP (processes #2013/50421-2; #2020/01779-5; #2021/08322-3; #2021/08534-0; #2021/10195-0; #2021/10639-5; #2022/10760-1) and National Council for Scientific and Technological Development - CNPq (processes #442147/2020-1; #440145/2022-8; #402765/2021-4; #313016/2021-6; #440145/2022-8), and São Paulo State University - UNESP for their financial support. This study is also part of the Center for Research on Biodiversity Dynamics and Climate Change, financed by the Sao Paulo Research Foundation - FAPESP. We also want to thank our anonymous reviewers for their observations and comments, which helped improve this work.
The following is Supplementary data to this article:

