





Planting of native trees has been adopted in many tropical regions worldwide as a central forest restoration method, but little is known concerning the role that these planted species play in catalyzing forest regeneration beneath their canopies. We investigated the role of animal-dispersed tree species in catalyzing the regeneration of woody species in the understory of restoration plantings. We assessed both the density and richness of tree seedlings within plots located beneath the canopy of both animal-dispersed and abiotic-dispersed tree species planted in three riparian forest restoration sites with ages of five, six and eight years. The proportion of animal-dispersed tree seedlings increased with plantation age. The richness of animal-dispersed tree seedlings was higher beneath animal-dispersed trees in the eight-year-old planting. The density of animal-dispersed tree seedlings was higher under animal-dispersed trees when sites were analyzed altogether. The top three species in regeneration density beneath the canopy were animal-dispersed trees, and from the top ten, seven were animal-dispersed species. We suggest that animal-dispersed pioneer trees which facilitate natural regeneration and promote a high density and richness of woody species beneath their canopies should be considered as “framework” species for tropical forest restoration.
Historical land uses in many human-modified tropical landscapes have compromised the resilience of natural ecosystems, and thus hampered their potential for self-recovery after abandonment (Chazdon, 2003). In order to recreate the conditions necessary for secondary succession in such a scenario, active restoration is needed to help overcome limitations concerning seed-dispersal and micro-site conditions (Holl and Aide, 2011). The planting of native trees has thus been adopted in many tropical regions worldwide as one of the main methods for restoring agricultural lands in this context (Rodrigues et al., 2011). Little is known however about the role that the species used in those plantings play in catalyzing forest regeneration beneath their canopies. Assessing the differing performances of planted tree species in permitting the spontaneous regeneration of other native species is thus a key tool for improving the ecological efficiency of restoration plantings.
Different approaches have been adopted toward the selection of tree species. They are usually based on the assumed functional role of the species in recovering degraded lands, and follow classifications such as: pioneer and non-pioneer species, filling and diversity species (Rodrigues et al., 2009), nurse plants (Padilla and Pugnaire, 2006), and “framework” species (Blakesley et al., 2002). Most of these approaches have focused on the reestablishment of a forest structure that is able to facilitate succession, but the role of such species in enhancing seed arrival remains little studied.
The reintroduction of animal-dispersed tree species in degraded lands may be particularly important for forest restoration in highly fragmented landscapes, since they can contribute to overcome dispersal limitation by attracting seed dispersers that may bring in their guts seeds they have consumed nearby (Lindell et al., 2012). However, animal-dispersed species do not form a homogeneous functional group, and may contain species with a distinct ability to attract frugivorous birds and bats to restoration sites, as a result of fruiting phenology, and fruit yield, size, nutritional value, smell and color (Wunderle, 1997). Consequently, outcomes for plant regeneration in restoration sites can differ widely amongst animal-dispersed species, and the identification and utilization of plant species that are more attractive to frugivores may enhance restoration success. An assessment of the regeneration community beneath the canopy of planted tree species may thus be useful for indicating which species should be favored in restoration plantings (Wydhayagarn et al., 2009). In this sense, we sought to investigate the role of animal-dispersed tree species in catalyzing the regeneration of woody species in the understory of restoration plantings. We aimed to answer the following questions: (1) is the richness and density of animal-dispersed tree seedlings higher under the canopy of animal-dispersed planted trees than under the canopy of abiotic-dispersed ones? (2) Is the regeneration of exotic animal-dispersed species higher under the canopy of animal-dispersed planted species? (3) Which planted tree species have a higher density of animal-dispersed tree seedlings under their canopies?
Material and methodsStudy sitesThe study was carried in Piracicaba, São Paulo State, southeastern Brazil, in a region originally covered by Seasonally Tropical Dry Forest, within the Atlantic Forest biome (22°42′S, 47°37′W; altitude: 546m; climate: Cwa, Koeppen's system). The study sites exist within a highly fragmented landscape with less than 10% forest cover that is principally a matrix of sugar-cane fields and urban zones. We selected three riparian forest restoration plantings with ages of five, six and eight years; the first two sites were previously occupied by pastures of the African grasses Urochloa spp., and the eight-year-old site by annual crops. Restoration planting was carried out in all sites using approximately 80 native tree species, which were planted in a 3×2m spacing scheme (1.667 seedlings per ha) and stimulated by the control of exotic grasses (usually Urochloa spp. and Panicum maximum) via mechanical mowing and herbicide application. Bird communities in these plantings are dominated by generalist species, typical of disturbed sites (Alexandrino et al., 2013).
Data collectingWe counted and identified to species every tree seedling taller than 10cm present in 2×3m plots, these plots being located under the canopy of both animal-dispersed and abiotic-dispersed trees planted in each restoration site. The trunk of the tree was located at the center of each plot. We classified each surveyed tree seedling into three categories, each category consisting of two classes: species origin (native or exotic), colonizing status (local or immigrant) and dispersal syndrome (animal-dispersed or abiotic-dispersed species). We considered as native the seedlings belonging to species that are known to occur naturally in the study region and as exotic all those belonging to species that do not naturally occur in the study region. We classified as local the seedlings belonging to planted species and as immigrant those that do not belong to any of the tree species planted at the specific restoration site where the seedling was surveyed. To better explore the results when performing analyses, we created new categories by combining two or three of the seedling categories previously mentioned.
We evaluated 215 planted trees, amounting to 86, 74 and 55 trees at the five, six and eight-year old sites respectively. Altogether, we assessed natural regeneration under the canopy of 21 planted tree species (four to six animal-dispersed species, and four to seven abiotic-dispersed species per restoration site), including pioneers and non-pioneers (Table S1). All field data was collected in July 2011.
Data analysisWe calculated tree seedling species richness and density under the canopy of planted trees, in terms of the absolute and proportional values according to the total number of tree seedlings. These calculations were performed separately for each restoration site and for each of the three seedling categories described above (origin, colonizing status and dispersal syndrome).
We used the non-parametric chi-square test to compare the proportions of animal-dispersed×abiotic-dispersed, local×immigrant, and native×exotic tree seedlings among restoration planting ages. These analyses were undertaken to infer whether there was a notable change in the tree seedling community according to the age of the restoration planting.
To compare the richness of animal-dispersed seedlings under the canopy of animal-dispersed and abiotic-dispersed planted trees we generated rarefaction curves, and this was due to the fact that in all restoration sites the proportion of planted trees investigated varied between the two dispersal syndromes. Rarefaction curves were obtained with 1000 simulations, separately for each site, using the EcoSim software (Entsminger, 2012). To compare the density of animal-dispersed tree seedlings under animal-dispersed versus abiotic-dispersed planted trees for each restoration planting age, and also the density of native and exotic animal-dispersed immigrant seedlings under the canopy of animal-dispersed versus abiotic-dispersed planted trees, we performed an unpaired Student's t-test. Finally, we calculated the density of animal-dispersed plants beneath the canopy of each planted species and presented the results graphically according to a rank of regeneration density. For all of the analyses described in this paragraph, we considered only tree seedlings belonging to immigrant (non-planted) species in order to avoid including plants that could have originated from trees planted in the restoration sites. In all cases, we employed a p critical value of 0.05.
ResultsWe found 94 different woody species under the canopy of the planted trees, 76 of these being native ones. The majority of tree seedlings belonged to immigrant, native and animal-dispersed species (Table 1). Altogether, we surveyed 2593 individual seedlings under the canopies of 215 planted trees (2.01plantsm−2); with 70.3% of these seedlings belonging to animal-dispersed species and 45.5% of them being animal-dispersed seedlings of immigrant species. The proportion of animal-dispersed and abiotic-dispersed tree seedlings varied according to plantation age (χ2=39.01; df=2; p<0.001), with an increase in the proportion of animal-dispersed with planting age (from five to six years old χ2=17.33; df=1 and p<0.001; from six to eight years old χ2=4.23; df=1 and p=0.04) (Table 1). A similar pattern of variation in the proportion of animal-dispersed seedlings with plantation age was found when only immigrant species were included in the analysis (χ2=23.39; df=2 and p<0.001). In this case, we found a higher proportion of animal-dispersed seedlings within the eight-year-old site in comparison to the younger restoration sites (from five to eight years old χ2=19.18; df=1 and p<0.001; from six to eight years old χ2=19.17; df=1 and p<0.001).
Richness and density of the tree seedling community regenerating in five, six and eight years old restoration plantings (Piracicaba-SP, Southeastern Brazil). Values between parentheses are the percentage of total number of species or density in each restoration planting.
| Group | Species richness | Seedling density (plantsm−2) | |||||
|---|---|---|---|---|---|---|---|
| 5y | 6y | 8y | 5y | 6y | 8y | ||
| Dispersal syndrome | Abiotic-dispersed | 19 (35%) | 25 (43%) | 22 (36%) | 0.80 (36%) | 0.52 (27%) | 0.39 (22%) |
| Animal-dispersed | 35 (65%) | 33 (57%) | 39 (64%) | 1.44 (64%) | 1.40 (73%) | 1.39 (78%) | |
| Origin | Immigrant species | 32 (59%) | 32 (55%) | 34 (56%) | 1.07 (48%) | 0.98 (51%) | 1.27 (71%) |
| Local species | 22 (41%) | 26 (45%) | 27 (44%) | 1.17 (52%) | 0.94 (49%) | 0.52 (29%) | |
| Identity | Exotic | 9 (17%) | 14 (24%) | 8 (13%) | 0.67 (30%) | 0.44 (23%) | 0.35 (20%) |
| Native | 45 (83%) | 44 (76%) | 53 (87%) | 1.57 (70%) | 1.48 (77%) | 1.43 (80%) | |
| Total | 54 (100%) | 58 (100%) | 61 (100%) | 2.23 (100%) | 1.92 (100%) | 1.78 (100%) | |
The proportion of local and immigrant tree seedlings varied among restoration sites (χ2=91.01; df=2; p<0.001). We found a higher proportion of immigrant species in the eight-year-old site (from five to eight years old χ2=86.28; df=1 and p<0.001; from six to eight years old χ2=59.11; df=1 and p<0.001) (Table 1).
Estimated richness for the tree seedling community did not significantly differ between dispersal syndromes within the two younger restoration sites (Fig. 1A and B). There was however a tendency of higher richness of animal-dispersed species in the eight-year-old plantation, revealed after the sample of 25 planted trees (Fig. 1C). The density of animal-dispersed tree seedlings did not differ between planted trees dispersal syndromes for each individual age, but it was higher under animal-dispersed trees when sites were analyzed altogether (Fig. 2A).
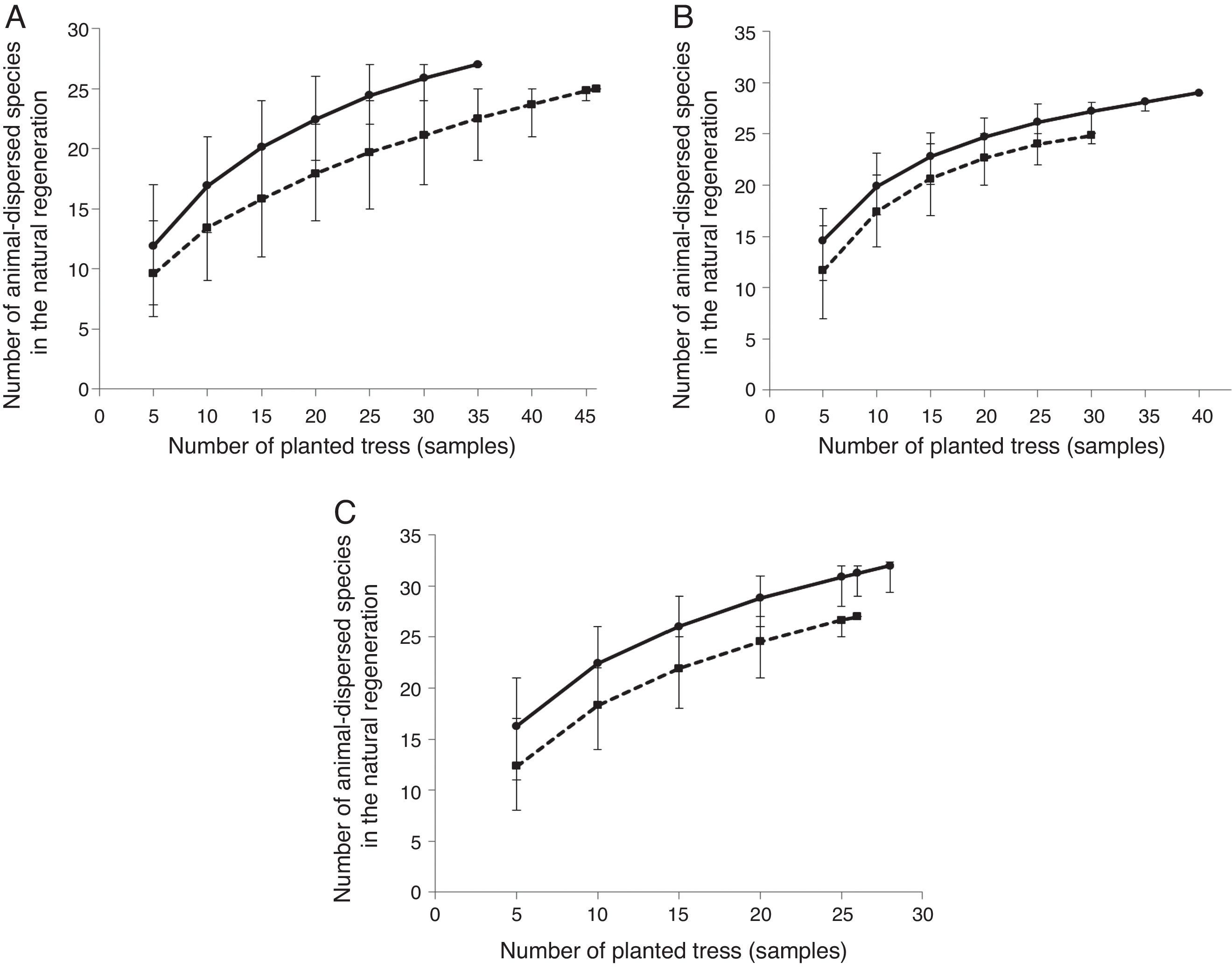
Number of animal-dispersed tree species under the canopy of abiotic-dispersed (dashed line) and animal-dispersed (continuous line) planted trees in five (A), six (B) and eight-years-old forest restoration plantings (C). Only seedlings of immigrant (non-planted) native species were accounted. Rarefaction curves were obtained with 1000 simulations. Vertical bars represent the 95% confidence intervals.
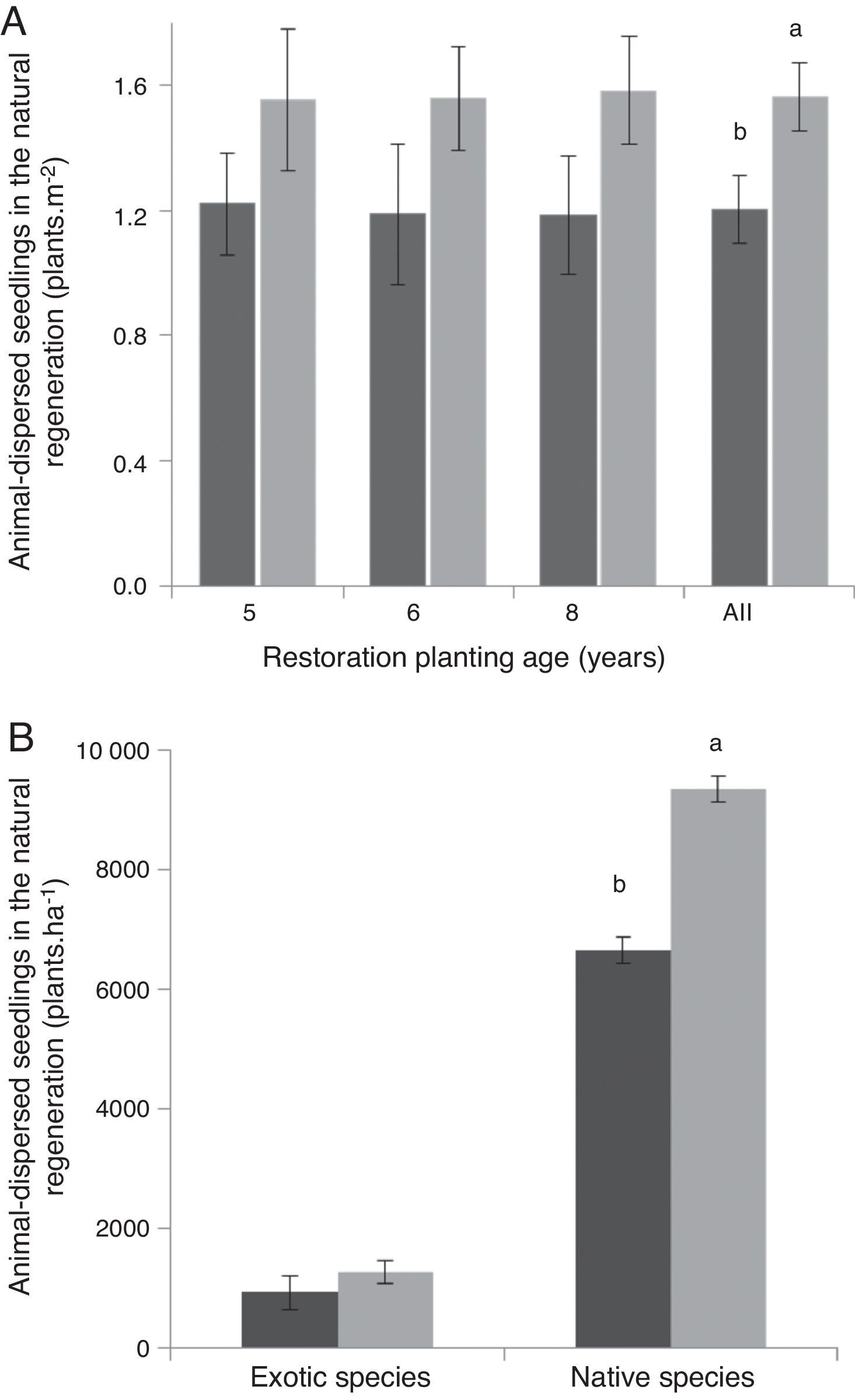
Mean regeneration density (±standard error) of animal-dispersed seedlings in forest restoration plantings of different ages (A) and of exotic and native animal-dispersed species (B) under the canopy of animal-dispersed (light gray) and abiotic-dispersed trees (dark gray) in Atlantic Forest restoration plantings, Piracicaba, southeastern Brazil. Only immigrant species (non-planted ones) were accounted. Bars followed by different letters significantly differ in the density of animal-dispersed seedlings (Student's t-test, p<0.05).
We found 18 exotic tree species present under the canopy of the planted trees, six animal- and 12 abiotic-dispersed species. Although the density of native tree seedlings was higher under the canopy of animal-dispersed trees, we found no difference in the density of exotic animal-dispersed immigrant seedlings connected to the dispersal syndrome (animal-dispersed or abiotic-dispersed) of the planted tree (Fig. 2B). In addition, the proportion of native and exotic seedlings under the canopy did not differ between animal- and abiotic-dispersed planted trees (χ2=0.035; df=1 and p=0.8524).
The mean regeneration density of animal-dispersed species under the canopy of the planted species varied from 3095plantsha−1 for Luehea divaricata (wind dispersed) up to 16,429plantsha−1 for Trema micrantha (animal dispersed), giving rise to a variation of 530% in the regeneration density. The top three species in regeneration density were animal-dispersed trees, and from the top ten, seven were animal-dispersed species (Fig. 3).
Discussion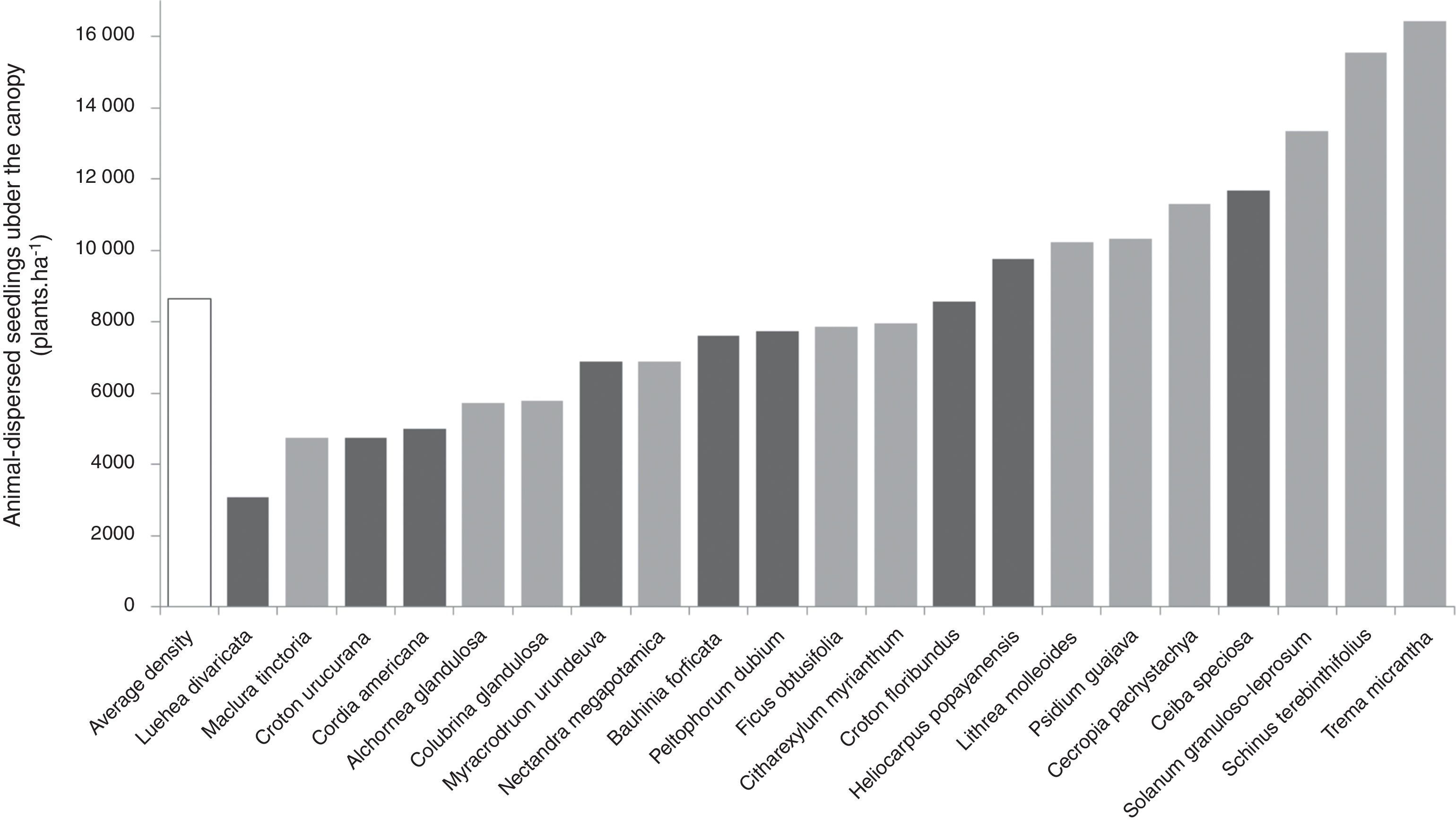
Despite only evaluating relatively young restoration plantings immersed in a highly fragmented landscape, we found high richness and density of animal-dispersed immigrant tree species regenerating under the planted trees. These restoration plantations thus appear to be actively functioning as successful catalysts of natural regeneration in degraded areas situated within a region assumed to contain strong limitations to dispersal, as indicated by the high establishment of immigrant animal-dispersed species from adjacent areas to the forest restoration sites. Since most of the species and individuals regenerating under the canopy of the trees in the restoration plantations belonged to immigrant (non-planted) species, our results suggest that in a relative short period of time, the composition of the plant community under restoration will be considerably transformed from the planted one. Furthermore, our results show that the planted tree species do not have, at least during the initial years after planting, a major role as seed and seedling sources for forest regeneration.
Higher natural regeneration richness under animal-dispersed trees in the eight-year-old site and the similarity between the two younger restoration sites may be a consequence of an increase in fruit abundance in the older site. This increase may be due to a greater fruit yield and/or number of animal-dispersed trees that matured to the reproductive stage in the older site, whilst in the younger plantations fruit production must not have yet been sufficient to produce a difference in the performance of animal-dispersed trees in comparison to the abiotic-dispersed sites (Wunderle, 1997). Although initially attracted into restoration zones by animal-dispersed trees, frugivores do not disperse seeds only under those trees (Wunderle, 1997); they may use any kind of tree as feeding or resting perches, dispersing seeds via defecation and regurgitation. Indeed, we did find a relatively high density and richness of animal-dispersed seedlings regenerating beneath the canopies of abiotic-dispersed trees. Additionally, the majority of potential seed dispersers found in the restoration sites studied is not strictly frugivores (Alexandrino et al., 2013) and may use any type of tree for foraging insects and other food sources, thus defecating or regurgitating seeds throughout the forest.
These seed dispersers – typical of disturbed areas – could also be playing an undesirable role in disseminating animal-dispersed exotic species. However, we did not find that exotic tree seedlings are more abundant under animal-dispersed trees than abiotic-dispersed, neither did we find any difference in the proportion of exotic and native tree seedlings connected to the dispersal syndrome of the planted trees. Therefore, in comparison with native trees, exotic ones are not particularly likely to be favored under the canopy of animal-dispersed trees. Nevertheless, it remains necessary to improve our understanding of the interaction between fauna and animal-dispersed exotic trees in order to prevent future biological invasion processes, and all of the subsequent costs that the management of exotic species in forest restoration sites entails (Buchley et al., 2006).
In spite of the overall patterns observed for animal- and abiotic-dispersed trees, great variation existed amongst species performance in catalyzing the understory regeneration. We found great variation in the density of tree seedlings under the canopies of the 21 investigated species. This is consistent with other studies (Fink et al., 2009) and was in fact expected since there is significant variation in the traits of animal-dispersed tree species concerning frugivore attraction; for example fruit color, smell, size and phenology (Wunderle, 1997). A key category of species that help to facilitate the regeneration of woody species are animal-dispersed pioneer trees. Pioneer trees are frequently used in tropical forest restoration plantations (Souza and Batista, 2004; Rodrigues et al., 2011) as they grow fast and tolerate competition with invasive grasses (Campoe et al., 2014). Due to the importance of fauna on catalyzing regeneration in forest restoration plantings, we suggest that the potential to attract seed-dispersing animals be a key criterion in the selection of tree species for forest restoration projects. Following from this conclusion, we recommend animal-dispersed pioneer trees be used as “framework” species for tropical forest restoration (Blakesley et al., 2002).
Concluding remarks and further advancesWe conclude that animal-dispersed trees, especially pioneers, enhance early regeneration in the studied forest restoration sites. We suggest they should be considered top priorities for tropical forest restoration. In addition, we recommend further studies to address knowledge gaps regarding the role that seed dispersal plays in catalyzing secondary succession in forest restoration plantings. Firstly, it is important to better investigate which animals consume the fruits of each planted tree species, so that restoration practitioners may select tree species that are especially attractive to seed dispersers (Lindell et al., 2012). In addition, research concerning the reproductive phenology of trees within restoration sites should be pursued and should relate to the supply of resources to animals.
Conflicts of interestThe authors declare no conflicts of interest.
The following are the supplementary data to this article:


