









Modifications in the structure of grasslands have been responsible for the decline of several avian species due to a reduced quality in available habitat. Currently, applications of the herbicide glyphosate have been introduced to promote winter forage production in the Flooding Pampas of Argentina. Rotational grazing has been another technique used to promote winter forage production. Here, we evaluated different grasslands managements in cattle production systems on species richness, composition, and abundance of bird assemblages in grasslands Pampas. We put emphasis on responses of two neartic migratory shorebirds, the Buff-breasted Sandpiper (Calidris subruficollis) and the American Golden Plover (Pluvialis dominica). The three treatments on natural grassland were: (1) paddocks of Ryegrass growth promotion through the application of herbicide glyphosate; (2) paddocks managed with Rotational grazing, and 3) paddocks managed with Continuous grazing. We considered three periods of the breeding cycle of the birds (early, late, post-breeding) for two years (2011/2012, 2012/2013). Vegetation cover lower than 10 cm was greater in both Rotational and Continuous grazing sites in comparison to the ryegrass promotion sites. A NMDS analysis revealed a clear separation in structure of bird communities among the three grazing treatments. The abundance of total birds and of short grassland birds was lower in grasslands with Ryegrass promotion. An indicator species analysis revealed the associations of shorebirds with Rotational and Continuous grazing sites. Grassland management with glyphosate application had an effect on the structure of assemblages of grassland birds in the Flooding Pampas, which was negative particularly with Nearctic migrants of conservation importance.
During the last decades, concerns about the environmental impact and sustainability of beef production have led to calls to revise these production systems worldwide (Steinfeld et al., 2006; Eisler et al., 2014; Herrero et al., 2015). Beef production systems are quite diverse, as they are the result of the combination of local environmental characteristics, culture, and economies (Seré and Steinfeld, 1996). They can be grouped into technologies based on the use of inputs (herbicides, fertilizers, seeds, animal feed, etc.), and those that are based on knowledge of natural processes (primary production of native habitat, decomposition, etc.) (Bilenca et al., 2018).
Herbicide applications have been widely used in Europe and the USA in order to manage vast areas of temperate grasslands or to replace them by crops (Milligan et al., 2003; Mau Crimmins, 2007). This practice produces changes in the structure and composition of the vegetation and has been responsible for the decline of several species of birds (McLaughlin and Mineau, 1995). Birds are highly vagile and, as a consequence of herbicide application, some studies have shown decline of different species of birds through reduced quality of available habitat (see Azpiroz et al., 2012), which affects their exploitation of food, nesting sites and shelter.
The Pampas and Campos of southeastern South America are dominated by grassland ecosystems that extend from southern Brazil into Uruguay and central-eastern Argentina (Bilenca and Miñarro, 2004; Jahn et al., 2017). The region’s extraordinarily favorable conditions for agriculture have resulted in a drastic modification of its native vegetation since European settlement (León et al., 1984; Modernel et al., 2016). In Argentina, the Flooding Pampas is a natural grassland ecosystem characterized by short grasslands (including various species of Distichlis, Nasella, Piptochaetium) and tall grasslands (including various species of Bromus, Paspalum), and is the most important livestock region of the country (Soriano et al., 1991; Faverin et al., 2019). Pastoral landscapes in the Flooding Pampas are also the most extensive wildlife habitat in the intensive agricultural Pampas region and, therefore, represent key habitats for biodiversity (Bilenca and Miñarro, 2004; Codesido et al., 2013). However, continuous grazing has resulted in changes in composition and structure of vegetation that have been maintained until the present, as pointed out elsewhere for other temperate regions (Soriano et al., 1991; Modernel et al., 2016). As a result of intensive agricultural and continuous grazing, it has produced a replacement of grassland birds, highly dependent on tall grasses, by others that select short grasses (Isacch and Martínez, 2001; Codesido et al., 2012). Furthermore, Isacch and Cardoni (2011) have shown that Continuous grazing positively influences the abundance of two migratory shorebirds, the American Golden Plover (Pluvialis dominica) and the Buff-breasted Sandpiper (Calidris subruficollis) in certain grasslands near to the Atlantic coast of Buenos Aires province.
In recent years, agrochemicals, particularly the herbicide glyphosate, have been used in the native grasslands of the Flooding Pampas to promote the growth of winter Ryegrass Lolium multiflorum, a non-native annual species (Rodriguez and Jacobo, 2013). While Ryegrass production has increased, this practice reduces the coverage of perennial C3 grasses, promotes the establishment of stolonipherous grasses such as Cynodon dactylon, and affects the structure of grassland and its seed bank resulting in a deterioration in forage resources after a few years (Rodriguez and Jacobo, 2013). The limited data available has reported that avian communities and several grassland specialists have been negatively affected by certain grassland managements such as promotion of pastures by means of use of agrochemicals (Agra et al., 2015; Da Silva et al., 2015; Fontana et al., 2016). Until now we do not know the consequences of the application of glyphosate and Ryegrass promotion on the attributes of the bird assemblage of the native grasslands of the Flooding Pampas.
On the other hand, Rotational grazing has been shown to promote winter forage production of great value (Jacobo et al., 2000) by increasing species of high forage value and reducing bare soil through the accumulation of litter (Jacobo et al., 2006). While the economic advantage of such management has been shown (Deregibus et al., 1995), the effect of Rotational grazing on avian communities is not fully known. However, in some short salty grasslands near to the Atlantic coast, Rotational grazing has lower abundance of neartic migratory grassland shorebirds than Continuous grazing (Isacch and Cardoni, 2011).
Here, we present results of different grasslands managements in cattle production systems (Grassland treated with herbicide, Continuous and Rotational grazing) and their associations with avian assemblages in the Flooding Pampas of Argentina in terms of species richness, composition, and abundance. Grassland treated with the herbicide glyphosate increase winter forage production of Ryegrass, providing lower cover of short vegetation (<10 cm high) and higher cover of grass above 10 cm high. Then, we will expect that richness and abundance of short-grass birds will be lower in Ryegrass promotion sites with respect to Continuous and Rotational grazing sites. We put special emphasis on two Nearctic-Neotropical migrants (Jahn et al., 2017), which breed in the grasslands or arctic tundra of North America and overwinter in coastal areas of the Río de la Plata Grasslands (Lanctot et al., 2002; Isacch and Martínez, 2003; Aldabe et al., 2018), the near threatened Buff-breasted Sandpiper (Birdlife International, 2020a) and the American Golden Plover. Both insectivorous shorebirds associated to short-grasses of humid prairies and halophytic steppes (Isacch and Martínez, 2003; Lanctot et al., 2004) have been declared a global conservation priority due to loss of wintering habitat in South America, historical commercial hunting during migration, and ongoing population declines (US Shorebird Conservation Plan Partnership, 2016). In addition, the highest concentrations of shorebirds in the nonbreeding habitat appear to be in the coastal grasslands used for livestock production (Lanctot et al., 2002). At last, we analyze the results in a framework that helps to understand what type of grazing management reconciles the conservation of birds and the grasslands with beef production.
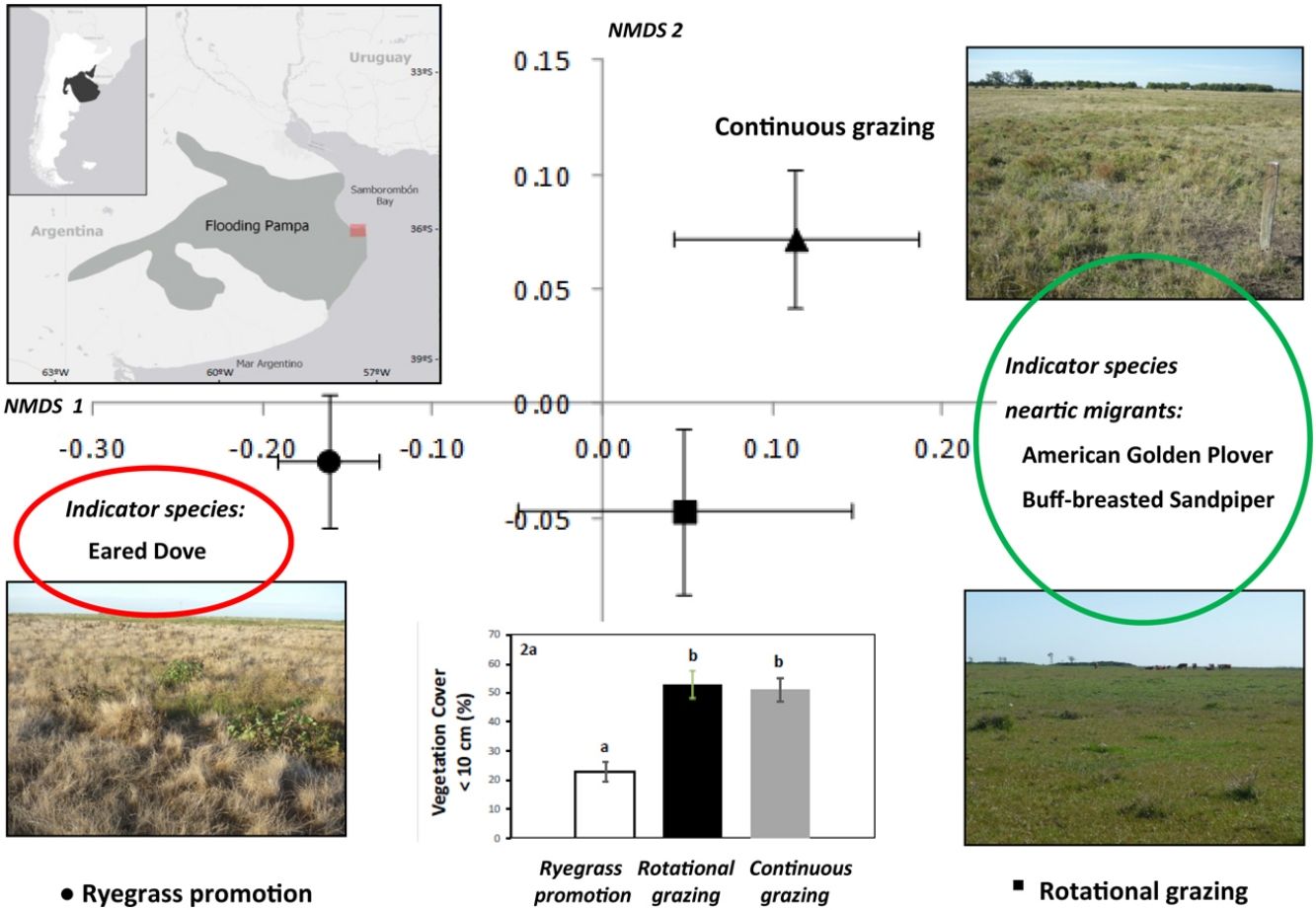
MethodsStudy areaOur study was conducted in the Pampas grasslands of southeastern South America (SESA grasslands; Azpiroz et al., 2012) extending along the coast of Samborombon Bay (Fig. 1) and covering about 244,000 ha. This region is temperate with hot summers (December–March) and cool, wet winters (June–August), with a mean annual temperature of 15 °C. The annual rainfall is around 1000 mm; the mean monthly rainfall varies between 50 and 150 mm. Except for small forest patches of Celtis ehrenbergiana, all other vegetation is mainly herbaceous. Vegetation is arranged in a complex mosaic of herbaceous communities mainly determined by edaphic factors (subtle topography and soil salinity gradient), while latitudinal (regional) variation plays a secondary role (Vervoorst, 1967). Grasslands have evolved under light grazing conditions but were heavily disturbed after the arrival of Europeans in the 16th century with the introduction of domestic herbivores and, probably, frequent fires (Soriano et al., 1991). The original community was characterized by a ~60 cm grassland dominated by tussock grasses including various species of Nasella, Piptochaetium, Bromus and Paspalum (Vervoorst, 1967) and in the lowland areas that are affected by temporary water supplies was characterized by Paspalidium paludivagum and Leersia hexandra. The study area is an important wintering site for migrant neartic birds (Lanctot et al., 2002; Birdlife International, 2020b) and is listed as an Important Bird Area (IBA) of Argentina due to several neotropical migrant species as well as endangered grassland species such as the Dot-winged Rail Porzana spiloptera (Birdlife International, 2020c) and the Bay-capped Wren-Spinetail Spartonoica maluroides (Birdlife International, 2020d).
At the top, location of the study area within the area of the Flooding Pampas, Argentina; yellow asterisks represent the fields surveyed. Source: 2020 Google Earth, Cnes/Spot Images. At the bottom, images of the three treatments within the Flooding Pampas of Argentina where birds and vegetation were sampled: ‘‘Ryegrass promotion” (left), ‘‘Rotational grazing (middle); ‘‘Continuous grazing’’ (right).
We conducted surveys on ranches located in General Lavalle County, Buenos Aires province, where cattle-breeding and farming under traditional continuous grazing are the main activities. Livestock density is typically 0.7–1 head of cattle ha–1. We defined three management practices (treatments, see Fig. 1): (1) Grassland treated with the herbicide glyphosate (hereafter, Ryegrass promotion) that regularly received late-summer applications to promote winter forage production (i.e., Ryegrass). Glyphosate was applied with terrestrial spray equipment at a dose of four to five l/ha in a single application during the first week in March each year. At this time, warm-season species are growing actively and cool-season species (Ryegrass) start their growing period (see Rodriguez and Jacobo, 2010). This practice increased winter forage production per hectare and allowed improvement of stocking rate and meat production (Bilello and Zeberio, 2002); (2) Rotational grazing, grassland in which cattle were rotated among paddocks every 15–30 days during spring and summer and every 3–4 months during autumn and winter (see Jacobo et al., 2006), (3) Continuous grazing, grassland in which grazing was maintained throughout the year within the same paddocks (i.e., the modal system in the study area; see González-Fischer and Bilenca, 2020).
Avian assemblages and vegetation characteristics were evaluated in the three treatments. Our field work included three periods of the breeding cycle of the birds, early (the sampling was made in October), late (December) and post-breeding (late February/March), along two years (2011/2012 and 2012/2013). For each treatment we located systematically 12–15 count points, one per paddock (paddock size varied from 12 to 25 ha), separated from each other by at least 150 m to ensure their independence and separated by the same distance from fences, drainages and floodplains to avoid the edge effect. Surveys were conducted during the 3 h after dawn (0615 ± 10–0915 ± 10) and the 3 h before sunset (1710 ± 10v–v1950 ± 10). The same observer (MC) sampled each point count and recorded all birds seen or heard within a 50-m radius of the survey point during a 5-min period (following Bibby et al., 2000) excepting individuals in flight (Codesido et al., 2013; Agra et al., 2015). We used a handheld GPS (error ±5 m) to measure distances. We conducted one survey at each treatment by period (i.e., 35–45 point-counts per sample, thus a total of six samples and 247 point-counts over the study period, resulting in a sampling effort of 1235 min).
We determined avian species richness and abundance for each survey point for both SESA birds (Azpiroz et al., 2012) and non-SESA birds. SESA birds were grouped as short-grass species, broad species or tall-grass species following the criteria of Azpiroz et al. (2012). We followed Remsen et al. (2020) for taxonomy.
In addition, we also made a visual estimation of vegetation height and cover within each 50-m radius survey point (Matteucci and Colma, 1982) after completing the count of birds at each point. We divided each 50-m radius survey point into four quadrants according to the cardinal points (North, East, South and West), taking a measurement in each quadrant and finally averaging them to obtain an estimate per each 50-m radius survey point. According to the use of short grasslands by shorebirds (Isacch and Cardoni, 2011; Aldabe et al., 2018) and grassland birds (Agra et al., 2015) we classified vegetation height using a graduated ruler in the following three height strata: lower than 10 cm tall, 10–24 cm, and ≥25 cm. Then, the dominant plant species were identified, grouped according to functional groups (Lavorel et al., 1997) and the coverage of each group was identified and estimated for each of the height strata, considering the whole 50-m radius. In this way, each 50-m radius survey point had a different percentage of each cover stratum which, in turn, changes over time. We used Roitman and Preliasco (2012) to assign the grasses and forbs species to the following functional groups: Annual winter grasses, Legumes, Creeping summer grasses, Perennial winter grasses, Graminoids, Erect summer grasses, Forbs, Grass weeds, Dicotyledon weeds, and Erect summer dicotyledons.
Statistical analysisAttributes of bird assemblages were analyzed by generalized linear mixed models (Zuur et al., 2009). The generalized linear mixed model provides a useful approach for analyzing repeated measures data by accounting for the lack of independence between observations that are repeated over time and to model more than one error term (Zuur et al., 2009). In this regard, each survey was considered a repeated measure. The response variables were bird species richness and abundance, and the explanatory variables were grazing management technique (3 levels: Ryegrass promotion, Rotational grazing and Continuous grazing) and period of the breeding season (3 levels: early, late and post-reproductive) as well as the interaction between both factors. Grazing management technique and period were fitted as fixed effects and survey point as a random term. Since the variance was much greater than the mean, both species richness and abundances of species data fitted negative binomial distribution and a log link function was used in all models (Zuur et al., 2009). Since no significant interannual variation was observed in the climatic variables, the year factor was not included in the model (the points between years were considered independent). Plots of residuals and normal probability were examined in order to evaluate assumptions of normality and homogeneity of variance by means of graphical validation tools (Zuur et al., 2009). The same analysis was used to compare vegetation cover (response variable) at different heights between grazing treatments. Vegetation cover data fitted beta distribution and a logit link function was used in all models (Zuur et al., 2009).
To identify bird species characteristic of each habitat under each management technique, we used an indicator-species analysis (De Cáceres and Legendre, 2009). This analysis calculates an indicator value for each species on the basis of its relative frequency and relative abundance in each category. Indicator values can range from 0 (no indication) to 100 (perfect indication). A perfect indication for a given habitat means that the species was recorded on all points within that habitat and was not observed in any other habitat. The function also allows evaluation of indicator species of both individual treatments and combination of treatments, as explained in De Cáceres (2013). We tested indicator values for significance with a Monte Carlo randomization procedure which compares the observed indicator values to alternative values calculated from the same data and randomly assigned to habitat type. We report only species whose indicator values are significant (P < 0.05).
Community composition were analyzed using Nonmetric Multidimensional Scaling (NMDS) with the Bray–Curtis index, we plotted the six samples for each grazing management type (3 different level of seasons by 2 years) in a multidimensional space defined by the total number of individuals of each bird species to graphically represent associations among samples.
Most of these analyses were implemented with R statistical software, Version 3.5.3 (R Development Core Team, 2019). For generalized linear mixed models we used glmmTMB package (Magnusson et al., 2017), and for indicator-species analysis we used indicspecies package (De Cáceres, 2013). For multidimensional scaling we used Past (Hammer et al., 2001).
ResultsVegetation structureThe structure of the vegetation was different between grazing management treatments (Fig. 2). Vegetation cover – less than 10 cm – was greater in both Rotational and Continuous grazing sites in comparison to the Ryegrass promotion sites (Generalized linear mixed model, treatment effect: Tukey test, P < 0.05; Fig. 2a). For this cover stratum in both Rotational grazing and Continuous grazing we found a greater number of species and functional groups than in Ryegrass promotion sites, including Legume species (Lotus sp., Trifolium sp. Melilotus sp.), and other creeping summer grasses (Paspalum sp., Stenotaprhum secundantum, Distichlis spicata) of forage interest (Table 1), whereas Ryegrass promotion sites had Ryegrass as the dominant species and to a lesser extent some grass and dicotyledon weeds (Table 1). Vegetation cover at the 11–24 cm stratum was greater in Ryegrass promotion sites in comparison to the Rotational grazing sites (Generalized linear mixed model, treatment effect: Tukey test, P < 0.05; Fig. 2b). For this cover stratum, the Ryegrass was again the dominant species in herbicide application sites, while this species in addition to legumes and creeping summer grasses were the dominant species for Rotational grazing and Continuous grazing (Table 1). Finally, vegetation cover – greater than 25 cm – was similar among treatments (P > 0.05; Fig. 2c). Ryegrass promotion sites had the Ryegrass as the dominant species (Table 1), while Rotational grazing and Continuous grazing have this species in addition to perennial winter grass (Nasella neesiana, Jarava sp.) and legumes (Table 1).
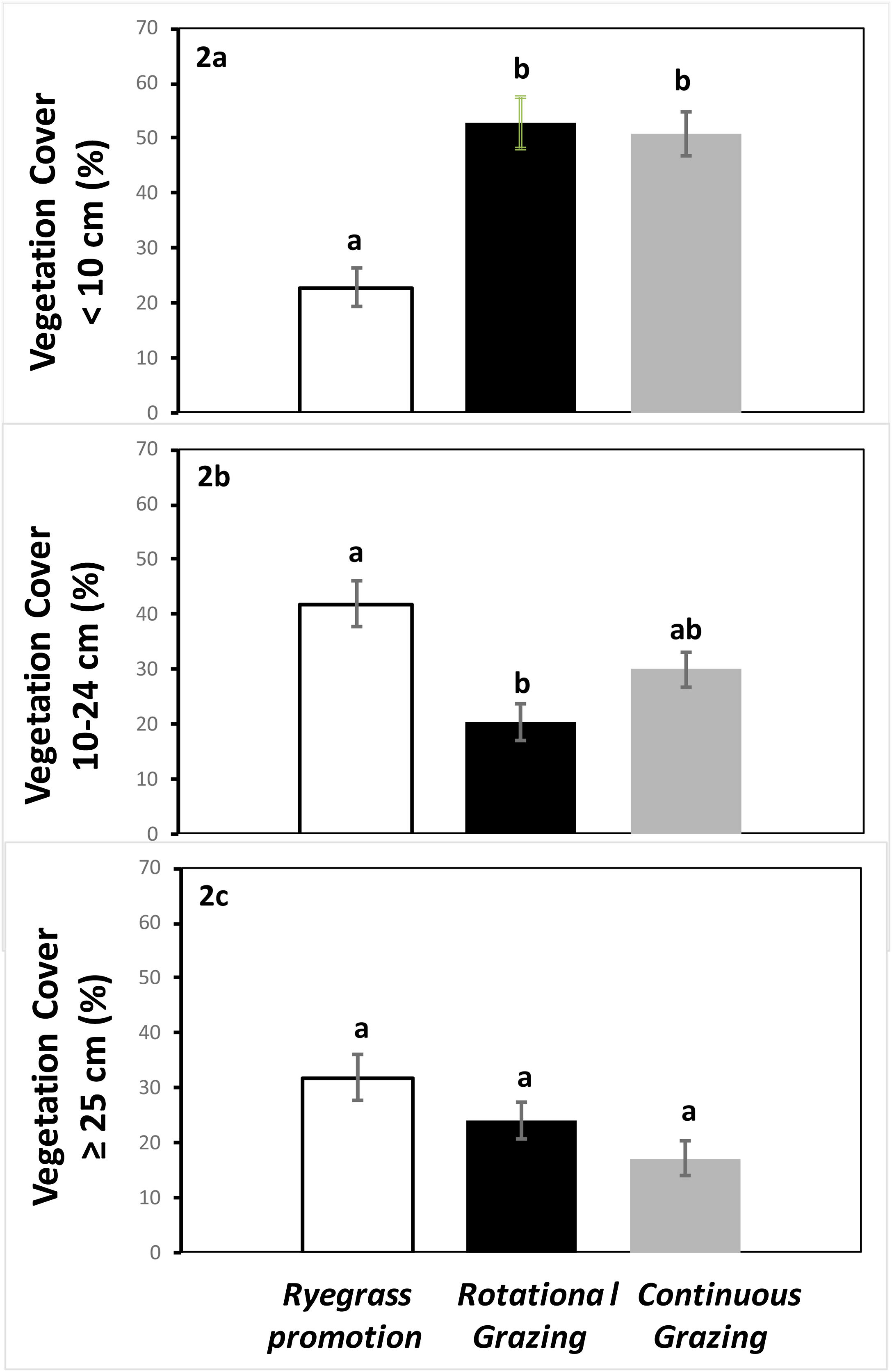
Total vegetation cover at different height strata for grasslands with Ryegrass promotion, grasslands with Rotational grazing and grasslands with Continuous grazing during spring-summer 2011/2012 and 2012/2013 in the Flooding Pampas, Argentina. Different letters indicate significant differences (Tukey test; P < 0.05).
Vegetation cover (%) of functional groups and species at different heights for treatments with Ryegrass promotion (RyP). Rotational grazing (RoG) and Continuous grazing (CoG) during spring-summer 2011–2012 and 2012–2013 in the Flooding Pampas. Argentina.
| <10 cm | 10–24 cm | ≥25 cm | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Functional groups | RyP | RoG | CoG | RyP | RoG | CoG | RyP | RoG | CoG |
| Annual winter grassesa | 79 | 20 | 15 | 67 | 31 | 24 | 45 | 25 | 19 |
| Legumesb | 26 | 11 | 2 | 22 | 2 | 2 | 12 | 3 | |
| Creeping summer grassesc | 5 | 33 | 49 | 14 | 30 | ||||
| Perennial winter grassesd | 2 | 1 | 5 | 7 | 3 | 24 | 28 | ||
| Graminoidse | 3 | 4 | 3 | ||||||
| Erect summer grassesf | 2 | 12 | 6 | 13 | |||||
| Forbsg | 9 | 12 | 6 | 20 | |||||
| Grass weedsh | 8 | 5 | |||||||
| Dicotyledon weedsi | 8 | 7 | 8 | 13 | 8 | 8 | 18 | 10 | 8 |
| Erect summer dicotyledonsj | 2 | 6 | 10 | 20 | 24 | ||||
In total, we recorded 3853 individuals and 41 bird species, from which 27 were SESA species (6 short-grass species, 21 broad species and none tall-grass species) and 14 non-SESA species (Table 2).
Mean abundance (individuals/point) of bird species recorded in short grasslands with different cattle grazing in the Flooding Pampas of Argentina sampled in spring-summer 2011–2012 and 2012–2013: Ryegrass promotion (RyP). Rotational grazing (RoG) and Continuous grazing (CoG). The species are ordered by their abundance. The “height” column follows the Southeastern South American (SESA) species’ classification according to Azpiroz et al. (2012). Migrant shorebirds are in bold.
| Species | RyP | RoG | CoG | Height |
|---|---|---|---|---|
| SESA species | ||||
| American Golden Plover Pluvialis dominica | 0.41 | 6.73 | 4.94 | Short |
| Southern Lapwing Vanellus chilensis | 3.48 | 3.43 | 2.60 | Short |
| Correndera Pipit Anthus correndera | 1.95 | 1.71 | 1.76 | Broad |
| Buff-breasted Sandpiper Calidris subruficollis | 0 | 1.32 | 1.06 | Short |
| Grassland Yellow-finch Sicalis luteola | 0.60 | 0.90 | 0.82 | Broad |
| Brown-and-yellow Marshbird Pseudoleistes virescens | 0.95 | 1.40 | 0.59 | Broad |
| Greater Rhea Rhea americana | 0.20 | 0.12 | 0.45 | Broad |
| Austral Negrito Lessonia rufa | 0.04 | 0.19 | 0.35 | Short |
| Chimango Caracara Milvago chimango | 0.27 | 0.29 | 0.18 | Broad |
| Spotted Nothura Nothura maculosa | 0.28 | 0.18 | 0.17 | Broad |
| White-rumped Swallow Tachycineta leucorrhoa | 0.01 | 0.02 | 0.10 | Broad |
| Shiny CowBird Molothrus bonaeriensis | 0 | 0 | 0.09 | Broad |
| Campo Flicker Colaptes campestris | 0 | 0.08 | 0.06 | Broad |
| Southern Caracara Caracara plancus | 0.09 | 0.11 | 0.06 | Broad |
| Fork-tailed Flycatcher Tyrannus savana | 0.16 | 0.10 | 0.06 | Broad |
| Buff-winged Cinclodes Cinclodes fuscus | 0 | 0.05 | 0.05 | Short |
| Spectacled Tyrant Hymenops perspicillatus | 0 | 0.02 | 0.03 | Broad |
| Brown-chested Martin Progne tapera | 0 | 0 | 0.03 | Broad |
| Burrowing Owl Athene cunicularia | 0.11 | 0.05 | 0.01 | Short |
| Long-winged Harrier Circus buffoni | 0.02 | 0.01 | 0.01 | Broad |
| Firewood-gatherer Anumbius anumbi | 0 | 0 | 0.01 | Broad |
| Hudson’s Canastero Asthenes hudsoni | 0.02 | 0 | 0.01 | Broad |
| White-browed Blackbird Sturnella superciliaris | 0.06 | 0.10 | 0.01 | Broad |
| Cattle Egret Bubulcus ibis | 0.56 | 0.79 | 0 | Broad |
| Cinereous Harrier Circus cinereus | 0.01 | 0.01 | 0 | Broad |
| Barn Swallow Hirundo rustica | 0.01 | 0.01 | 0 | Broad |
| Great Pampa-Finch Embernagra platensis | 0.01 | 0.02 | 0 | Broad |
| Non SESA species | ||||
| White-faced Ibis Plegadis chihi | 0.24 | 0.54 | 3.33 | |
| Monk Parakeet Myiopsitta monachus | 0.15 | 0.40 | 0.17 | |
| Southern Screamer Chauna torquata | 0.02 | 0.12 | 0.04 | |
| Rufous-collared Sparrow Zonotrichia capensis | 0.02 | 0.10 | 0.03 | |
| Hooded Siskin Spinus magellanicus | 0 | 0.01 | 0.03 | |
| Saffron Finch Sicalis flaveola | 0 | 0 | 0.01 | |
| Eared Dove Zenaida auriculata | 0.33 | 0.04 | 0.01 | |
| Glittering-bellied Emerald Chlorostilbon lucidus | 0 | 0 | 0.01 | |
| Great Kiskadee Pitangus sulphuratus | 0 | 0.01 | 0.01 | |
| Whistling Heron Syrigma sibilatrix | 0.04 | 0 | 0 | |
| Brown-hooded Gull Chroicocephalus maculipennis | 0.53 | 0.02 | 0 | |
| Lesser Yellowlegs Tringa flavipes | 0.16 | 0 | 0 | |
| Hudsonian Godwit Limosa haemastica | 0 | 0.10 | 0 | |
| Black-winged Stilt Himantopus melanurus | 0.19 | 0.01 | 0 |
We observed 29 species and 947 individuals in the Ryegrass promotion, 33 species and 1595 individuals under Rotational grazing, and 32 species and 1311 individuals under Continuous grazing (Table 2). In addition, 20 species that were detected in the Ryegrass promotion sites, 23 species detected in Rotational grazing sites and 23 species detected in the Continuous grazing sites are considered representative of southeastern South American grasslands. Among SESA species, the American Golden Plover and Buff-breasted Sandpiper, both nearctic migratory shorebirds, represented 31% of the total number of individuals seen. Other abundant SESA species included the Southern Lapwing (Vanellus chilensis) and Correndera Pipit (Anthus correndera), followed by the Brown-and-yellow Marshbird (Pseudoleistes virescens), Grassland Yellow-finch (Sicalis luteola), and the White-faced Ibis (Plegadis chihi) a non-SESA species (Table 2).
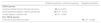
The NMDS showed a clear separation in structure of bird communities among the three grazing treatments (Fig. 3; NMDS stress = 0.19). The first ordination axis groups the Rotational and Continuous grazing sites and separated them from the Ryegrass promotion sites, while the second ordination axis separated Rotational grazing sites from Continuous grazing sites.
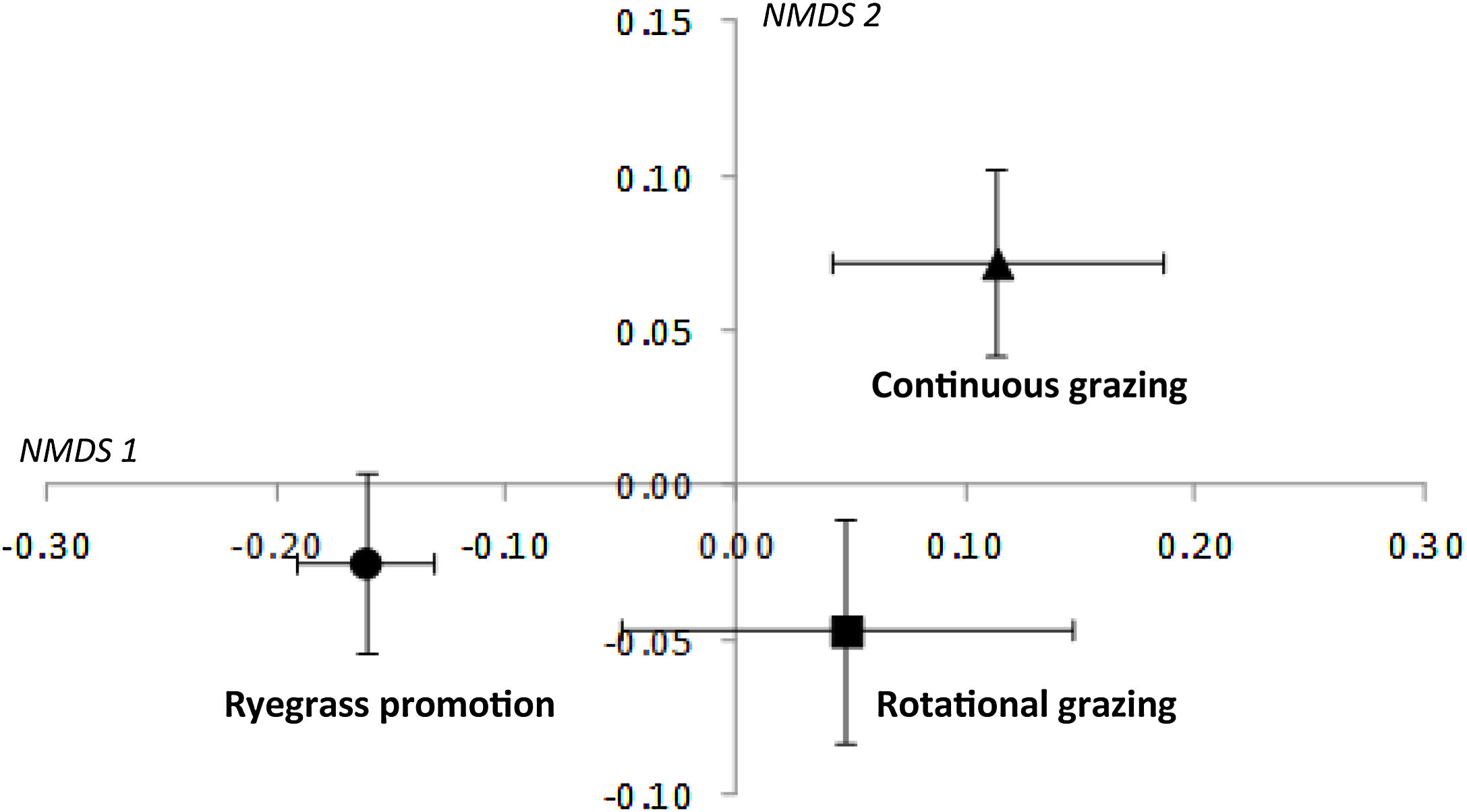
Nonmetric multidimensional scaling based on species’ abundance from 18 sample sites within three categories of grazing management (Ryegrass promotion, grasslands with Rotational grazing and grasslands with Continuous grazing) in the Flooding Pampas, Argentina. Symbols represent the mean and standard error of samples sites in each grazing treatment to enhance visual interpretation of the ordination (NMDS stress = 0.19).
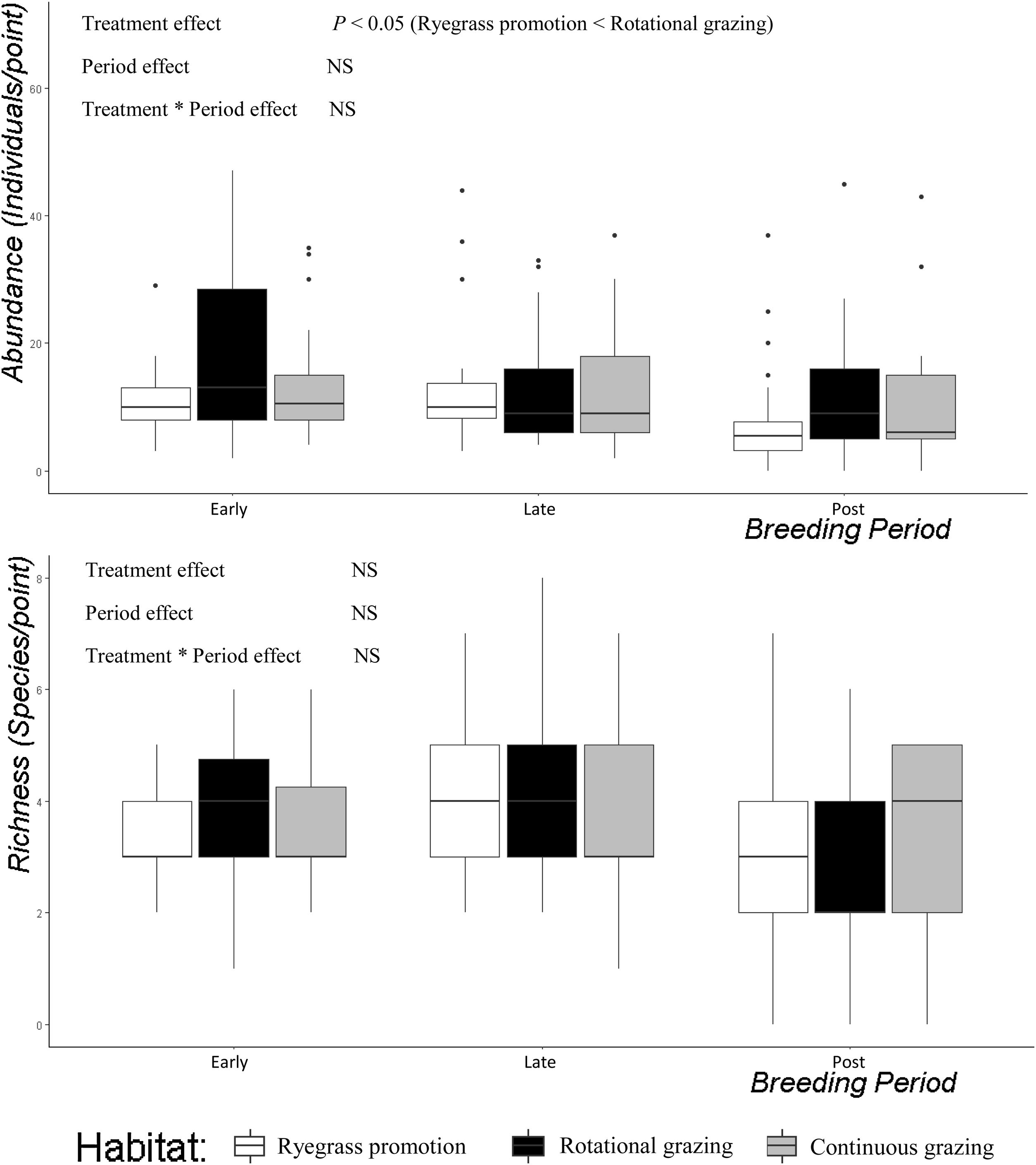
The total abundance of birds was lower in Ryegrass promotion sites in comparison with Rotational grazing sites (Generalized linear mixed model, treatment effect: Tukey test, P < 0.05; Fig. 4). In particular, the abundance of short-grass species was also lower in Ryegrass promotion sites when compared to Rotational and Continuous grazing sites (Generalized linear mixed model, treatment effect: Tukey test, P < 0.001 and Tukey test P < 0.01). The abundance of broad grassland species was similar among treatments (P > 0.05). Species richness was similar among treatments for all periods of the breeding cycle of the birds for all species (P > 0.05; Fig. 4), and for short and broad grassland birds (P > 0.05).
Indicator-species analysis identified four species with significant indicator values; three were associated with Rotational and Continuous grazing and one with Ryegrass promotion (Table 3). Three of the six species associated with short grasses are indicators for Rotational grazing and Continuous grazing, the American Golden Plover, Buff-breasted Sandpiper and Austral Negrito (Lessonia rufa), whereas indicator species for the Ryegrass promotion is the Eared Dove Zenaida auriculata, a non-SESA species (Table 3).
Indicator values (as percentage of perfect indication) and values of Monte Carlo test of significance of observed maximum indicator values. Only values significant with P < 0.05 are shown. SESA species: grassland birds in southeastern South America (Azpiroz et al., 2012).
| Rotational grazing + continuous grazing | Ryegrass promotion | |
|---|---|---|
| SESA species | ||
| American Golden Plover Pluvialis dominica | 90 (P < 0.001) | |
| Buff-breasted Sandpiper Calidris subruficollis | 36.7 (P = 0.031) | |
| Austral Negrito Lessonia rufa | 41 (P = 0.044) | |
| Non SESA species | ||
| Eared Dove Zenaida auriculata | 46.7 (P = 0.002) |
Overall, our results suggest that different cattle management practices affect abundance and structure of avian assemblages in grasslands of the Flooding Pampas. Our work, along with earlier studies (Isacch and Cardoni, 2011; Cardoni et al., 2015; Agra et al., 2015), broadens our understanding of the avian responses to grazing management practices in the southeastern South American grasslands (Azpiroz et al., 2012). Ryegrass promotion sites had lower total avian abundance than rotational grazing sites. Moreover, Rotational and Continuous grazing sites supported more abundances of short-grass species. Ryegrass promotion sites had a different vegetation structure, in particular a lower vegetation cover in the lowest stratum – less than10 cm – compared to Rotational and Continuous grazing sites. Herbicide application results in Ryegrass having the greatest coverage ratio and a lower representation of functional groups over the entire breeding bird period compared to Rotational and Continuous grazing sites. The responses observed in the characteristics of vegetation in the Ryegrass promotion sites is accordingly to reported in previous studies (Rodriguez and Jacobo, 2010).
Total abundance of birds was higher in Rotational grazing sites than in Ryegrass promotion sites. This result is consistent with changes reported in the Pampas grasslands of Rio Grande do Sul in Brasil, in which greater avian abundances were recorded in natural grassland sites compared to cultivated fields of Ryegrass (Fontana et al., 2016). A similar pattern were found in grasslands of central Europe (Batáry et al., 2007), and in North America (Murphy, 2003; Askins et al., 2007), in which greater avian abundances were recorded in extensive ranching sites compared to intensive agricultural areas.
Short-grass species had greater abundance in both Rotational and Continuous grazing sites over the entire breeding season in the natural grasslands. The pattern of bird abundance was greatly influenced by two migratory shorebirds, the American Golden Plover and Buff-breasted Sandpiper, which are indicator species of both grazing treatments. The arrival of shorebirds in early spring on the study area coincides with the beginning of the growing season of summer grasses. Shorebirds were absent from areas where grasslands had low percentages of short-grass cover (i.e. Ryegrass promotion sites), as were reported by previous studies (Lanctot et al., 2002; Isacch and Martínez, 2003; Isacch and Cardoni, 2011). In this regard, Aldabe et al. (2018) reported in the Campo grasslands from Uruguay that the American Golden Plover and Buff-breasted Sandpiper showed a relatively high probability of occurrence when grass height was between 2 cm and 5 cm and a very low probability of presence when grass height above 10 cm. The reasons behind this selection for short grass heights are unknown (Aldabe et al., 2018) but the absence of both species in the Ryegrass promotion sites in our study could be due to the low proportion of short grasses cover associated with an impoverishment of the functional groups of vegetation.
Other short grassland species recorded in the three grazing treatments were common and widely distributed, such as Southern Lapwing and Burrowing Owl Athene cunicularia (Narosky and Di Giacomo, 1993). Our observations support the natural history characteristics reported for these species in terms of foraging, cover, and nesting opportunities (Isacch and Cardoni, 2011; Dias et al., 2014; Agra et al., 2015; Azpiroz and Blake, 2016). A similar pattern was observed among grassland generalists, such as Greater Rhea, Correndera Pipit, Brown-and-yellow Marshbird, Grassland Yellow Finch, and Chimango Caracara (Agra et al., 2015; Azpiroz et al., 2012; Codesido and Bilenca, 2021; Isacch and Cardoni, 2011).
Among non-SESA species, the Eared Dove was the only indicator species recorded for Ryegrass promotion sites and which is also associated with cultivated and stubbled fields throughout the entire Pampas region (Codesido et al., 2015; Zufiaurre et al., 2019). In cropland landscapes, between the Pampas and the Espinal ecoregions of central Argentina, populations of the native Eared Dove have increased significantly in several provinces to such an extent that the species has been declared a pest (Bucher and Ranvaud, 2006). This species was noted to respond to Raygrass promotion in our study area, which is consistent since it is a granivorous species and as has been previously reported for agroecosystems of the Pampa ecoregion (Zufiaurre et al., 2017).
Grazing management and conservation implicationsDomestic European cattle have modified the floristic composition and structure of the grasslands of the Pampas (León et al., 1984). This process has drastically accelerated since the end of the 19th century, when cattle became confined and therefore Continuous grazing initiated (Deregibus et al., 1995). Traditional continuous grazing has deteriorated these grasslands, leading to 1) severe reduction of winter grasses which are highly preferred and severely defoliated during winter, 2) invasion by exotic and native forbs of low forage value, 3) soil denudation, and 4) reduction of stocking rate, animal performance, and net profitability (Deregibus et al., 1995).
Environmental and grazing management problems related to traditional continuous grazing have promoted the search for alternative management practices, including those based on the use of agrochemical inputs and simplification of the natural system versus those that are based on knowledge of natural processes (Bilenca et al., 2018). Our results are in agreement with management studies showing that agrochemical inputs may be more detrimental for wildlife conservation than those which are based on understanding ecological processes (Eisler et al., 2014; Bilenca et al., 2018, and references therein). Application of glyphosate in the native grasslands of the Flooding Pampa has been promoted to increase winter forage production. However, this practice changes species composition, floristic richness, diversity of seed bank and greatly reduces the summer and annual aboveground net primary production (Rodriguez and Jacobo, 2013; Rodriguez et al., 2018). The recurrent spraying of glyphosate in the Flooding Pampa grassland negatively affects ecosystem functioning and risks biodiversity loss in this natural habitat (Rodriguez et al., 2018). In particular, grassland management with the herbicide glyphosate has a negative effect on the functional groups of vegetation and in the structure of grassland bird assemblages in the Flooding Pampas.
Rotational grazing has usually resulted in an immediate increase of the stocking rate of about 30% (Jacobo et al., 2006). The increase of species of high forage value, decline in lower foraging value species, and the reduction of bare soil improve rangeland conditions and carrying capacity (Jacobo et al., 2006) due to the increase of winter forage productivity (Jacobo et al., 2000). Previous work in the study area has shown the potential of Rotational grazing to successfully attend both productive goals as well as conservation of endangered herbivores like Pampas deer Ozotocerus bezoarticus (Rodriguez et al., 2016), as well as conservation of the biodiversity of native birds (Vaccaro et al., 2020). In our study, shorebirds were closely associated with both Rotational and Continuous grazing management, whereas Ryegrass promotion using herbicides was detrimental for shorebirds.
Both livestock production and conservation compete for land in the Pampas ecoregion. Most wildlife biodiversity studies have focused on conservation rather than balancing biodiversity and domestic animal production, constraining the possibilities of benefiting both wild and domestic animals (Neilly et al., 2016; Martinez Ortiz et al., 2017). In this sense, conservation practices and livestock production in this region should be considered together, and discussed among farmers, conservationists, and government agencies (Dotta et al., 2015; González-Fischer and Bilenca, 2020).
FundingFinancial support was provided by the project “Grasslands and Savannas of the Southern Cone of South America: Initiatives for their Conservation” (GEF Project MSP Grant No. TF96757), FVSA, Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET, PIP 2010–2012 GI 11220090100231), and Universidad de Buenos Aires (UBACyT 2010-2012 GC20020090100070).
Ethics statementWe followed the protocol of the University of Buenos Aires, Argentina.
Author contributionsM.C. and D.B. equally conceived the idea, and design; M.C. collected the data and conducted the research; M.C. and D.B. wrote the paper; M.C. developed and designed the methods; M.C. analyzed the data; and M.C. and D.B. contributed substantial materials, resources, or funding.
Declaration of interestsThe authors (Mariano Codesido and David Bilenca) declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
We are grateful to the owners and workers of Estancias El Raigón, San Benito, La Gloria, Los Overos and El Divisadero for providing permission to work in their properties. Mario Beade from the Administración de Parques Nacionales (APN) provided housing and facilitated valuable logistic support. Fernando Miñarro and Pablo Preliasco from the Fundación Vida Silvestre Argentina (FVSA) helped us with logictic support and provided valuable information on grazing management. We appreciate the improvements in English usage made by Peter Lowther through the Association of Field Ornithologists' program of editorial assistance. The editor and three anonymous reviewers provided careful edits and very useful comments that enhanced this paper.

