



Spatial planning and decision-support tools facilitate the consideration of socioecological tradeoffs associated with extractive activities, but insufficient data resources often limit their application. Focusing on birds and mining concessions in the Northern Andes, we illustrate how publicly-available data can be used in spatial prioritization tools to identify where concessions have the potential to impact 22 species of Neotropical migratory birds. Concessions covered 11% of the land area of Colombia, Ecuador, and Peru, and disproportionately occurred within important areas for migratory birds in Colombia and Ecuador, but not Peru. eBird data showed that one-quarter of avian species, including the Olive-sided Flycatcher, Cerulean Warbler, and Canada Warbler, had >10% of their global populations on concession land during the non-breeding season. More worrisome, species of greater conservation importance and with larger population declines (1974–2014) were most likely to co-occur with mining. Our approach highlights how public biodiversity data can be used to predict, avoid, or mitigate potential ecological impacts from extractive activities.
Resource demands to support a burgeoning human population continue to accelerate rates of land conversion and threaten biodiversity and ecosystems around the world. Systematic conservation planning is one useful framework that identifies and prioritizes locations for conservation action based upon project objectives, ecological benefits, costs, risk, and other relevant information (Margules and Pressey, 2000; Bower et al., 2018; Schwartz et al., 2018). Although most prioritizations utilize species data (Sinclair et al., 2018), the resolution and quality of datasets vary widely and, thus, often have limited application (Cook et al., 2017). Fortunately, as large-scale and open-access data become more widely available (Sullivan et al., 2014), decision-makers can more precisely identify ecologically important areas and estimate impacts from proposed actions. In this research letter, we use migratory birds to illustrate how publicly-available data can facilitate strategic planning of mines and conservation in the Northern Andes.
More than half of migratory bird species in the world have declined over the past 30 years (Kirby et al., 2008), but conservation is complicated by the need to both work across geopolitical boundaries and support local peoples within biodiversity hotspots. Protected areas alone are not the solution. Not only do one-third of protected areas face intense human pressure (Jones et al., 2018), but only 9% of migratory bird species compared to 45% of nonmigratory species have their ranges adequately protected (Runge et al., 2015). Effective conservation at scale, thus, requires approaches that can be applied to working landscapes where human communities also draw heavily from local resources.
Our study was inspired by a global surge in mining concessions granted, especially in the Andes of South America with the privatization and internationalization of mining industries in the 1990s and 2000s (Devenish and Gianella, 2012; Perreault, 2013; Brain, 2017). Mineral exploration in Ecuador rose from 3% to 13% of the country's land area since 2016, including >30% of the countries ‘protected forests’, 14% of its indigenous lands, and may impact hundreds of vulnerable, threatened, and endangered species (Roy et al., 2018). Likewise, Brazilian mines now impact about one-third of the collective ranges of endemic anuran and birds (de Castro Pena et al., 2017). Even worse, 75% of active mines and concessions around the world overlap with areas of high ecological value, with one-third falling within otherwise intact ecosystems (Miranda et al., 2003).
The effects of mining are expected to vary with stage of development, and cumulative environmental impacts due to pollution, built infrastructure, altered geomorphology, as well as direct and indirect effects of habitat loss, degradation, and fragmentation, can be difficult to estimate or predict (Bridge, 2004). Because concessions define the spatial extent of mining explorations, they typically exceed the size of active mines. Nevertheless, exploration requires road construction, remote camps, drilling or trenching, and seismic survey – all of which expose species and their habitats to illegal hunting, pollutants, disturbance, and other human activities. Case in point, tree cover in Myanmar declined by 6% in mining concessions over 5 years (Papworth et al., 2017). Potential impacts, such as these, underscore the importance of careful planning that aims to avoid, reduce, and/or mitigate possible environmental harm. Despite widespread agreement about their usefulness, planning efforts are often constrained by a paucity of data, lack of expertise with quantitative and spatial methodologies, or even a reticence to engage due to perceived difficulty of the process.
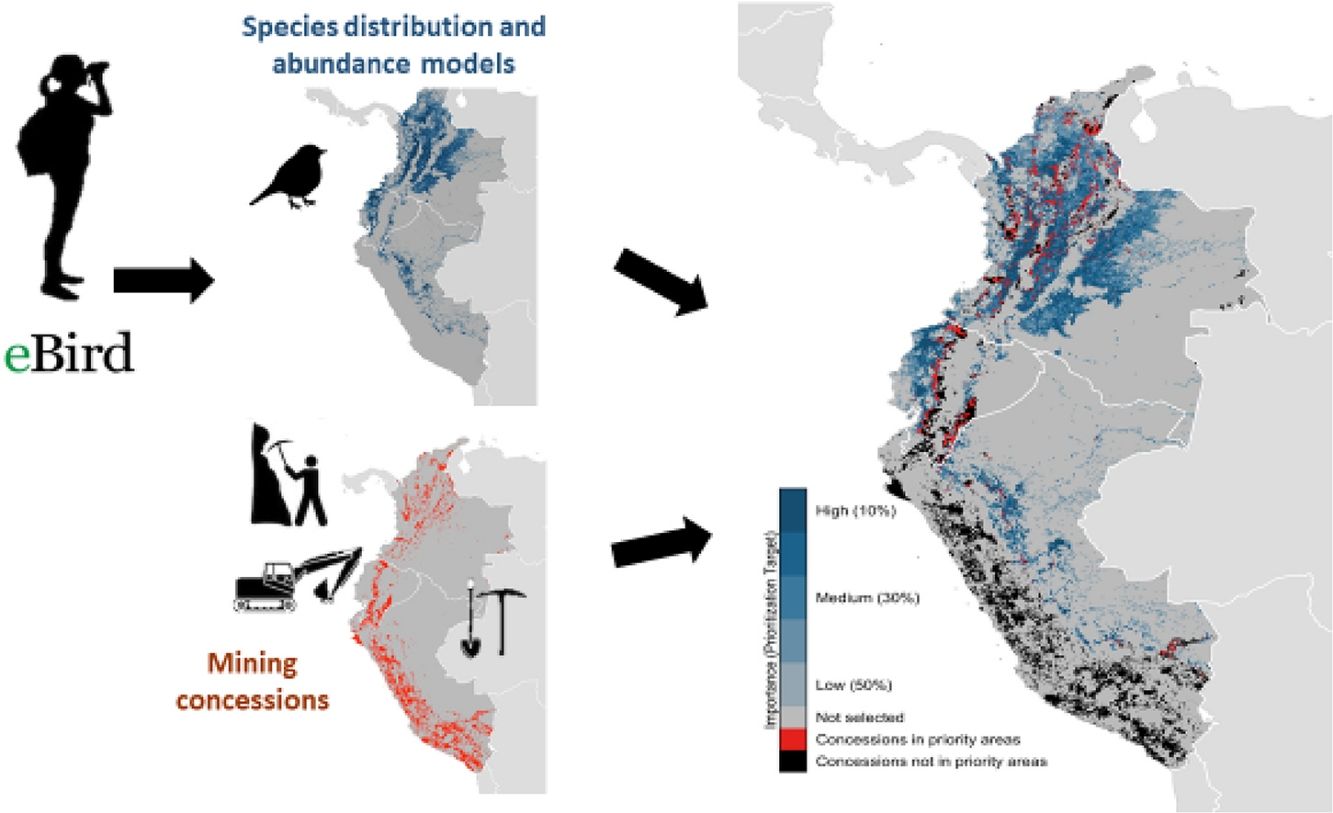
To illustrate the value of data-driven planning, we integrated data on mining concessions in northern South America with publicly-available data on avian distribution and abundance from eBird, a global platform at the Cornell Lab of Ornithology that collects millions of bird observations and checklists from volunteers (Sullivan et al., 2014). We then quantified the extent to which mine concessions overlapped key nonbreeding areas for 22 species of Neotropical migratory birds in Colombia, Ecuador, and Peru.
MethodsOur study focused on three countries in the Northern Andes–Colombia, Ecuador, and Peru – all countries that have experienced spikes in the establishment of mining concessions, and all recognized as important overwintering locations for many migratory birds. We based our assessments of bird species occurrence on distribution models developed using data from eBird, a citizen-science effort that represents the largest, most rapidly growing biodiversity database globally (Hochachka et al., 2012; Sullivan et al., 2014).
From bird count data in the 2016 eBird Reference Dataset (Fink et al., 2017), we estimated relative abundance across the Western Hemisphere for each week of the annual cycle using spatiotemporal exploratory models (STEM) (Fink et al., 2010; Johnston et al., 2015). These relative abundance estimates were made for each species over a uniform, equal-area grid of 8.4km×8.4km cells (hereafter “sites”). We then averaged the weekly abundance estimates from the non-breeding season (November to March) to produce a single map of estimated relative abundance for each species on their non-breeding grounds in the Northern Andes. Finally, these distributions were validated by scientists at the Cornell Lab of Ornithology. From an initial list of 237 species from the Western Hemisphere for which there were sufficient eBird data to reliably model distribution and abundance, we selected 22 that were (a) passerines, (b) migratory, and (c) distributed in the Northern Andes for at least part of their non-breeding stationary periods (Table 1).
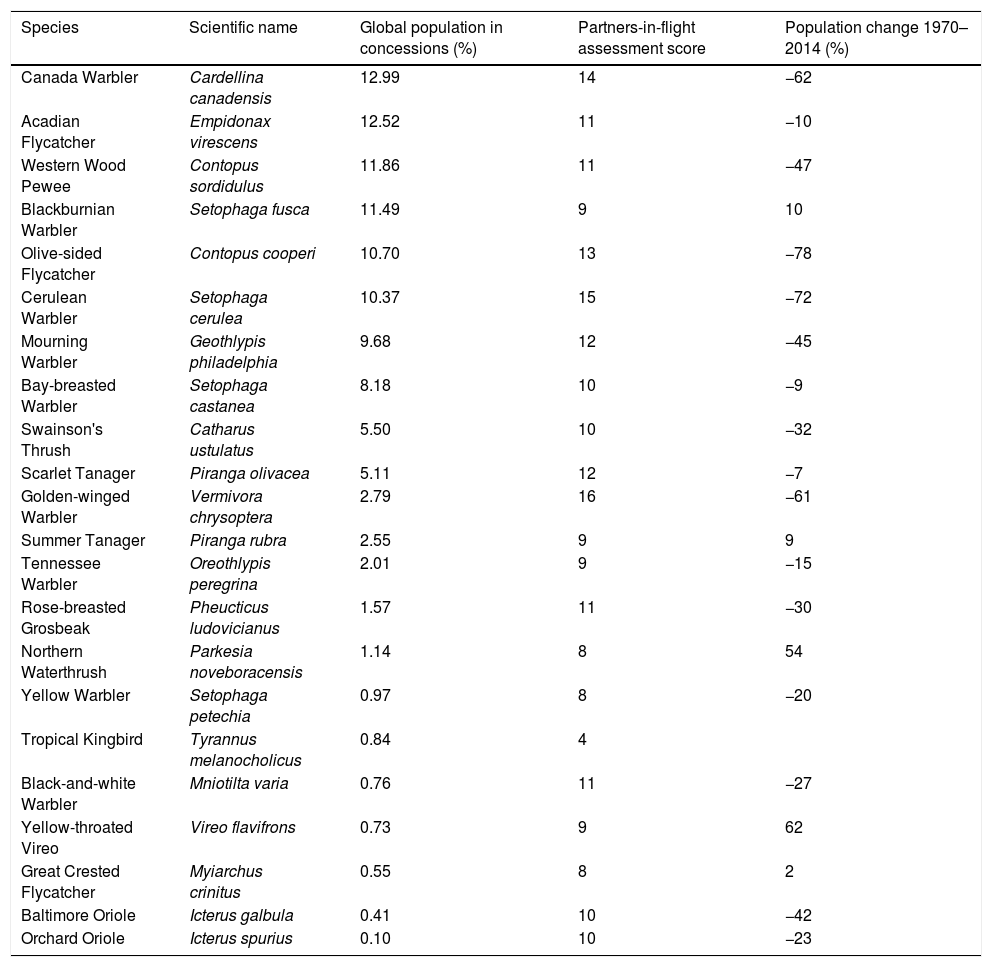
Percent global population of 22 of Neotropical migratory birds that falls within mining concessions in Colombia, Ecuador, and Peru based on eBird models, as well as the Partners-in-Flight assessment score (higher indicates greater level of conservation concern) and percent population change between 1970 and 2014.
| Species | Scientific name | Global population in concessions (%) | Partners-in-flight assessment score | Population change 1970–2014 (%) |
|---|---|---|---|---|
| Canada Warbler | Cardellina canadensis | 12.99 | 14 | −62 |
| Acadian Flycatcher | Empidonax virescens | 12.52 | 11 | −10 |
| Western Wood Pewee | Contopus sordidulus | 11.86 | 11 | −47 |
| Blackburnian Warbler | Setophaga fusca | 11.49 | 9 | 10 |
| Olive-sided Flycatcher | Contopus cooperi | 10.70 | 13 | −78 |
| Cerulean Warbler | Setophaga cerulea | 10.37 | 15 | −72 |
| Mourning Warbler | Geothlypis philadelphia | 9.68 | 12 | −45 |
| Bay-breasted Warbler | Setophaga castanea | 8.18 | 10 | −9 |
| Swainson's Thrush | Catharus ustulatus | 5.50 | 10 | −32 |
| Scarlet Tanager | Piranga olivacea | 5.11 | 12 | −7 |
| Golden-winged Warbler | Vermivora chrysoptera | 2.79 | 16 | −61 |
| Summer Tanager | Piranga rubra | 2.55 | 9 | 9 |
| Tennessee Warbler | Oreothlypis peregrina | 2.01 | 9 | −15 |
| Rose-breasted Grosbeak | Pheucticus ludovicianus | 1.57 | 11 | −30 |
| Northern Waterthrush | Parkesia noveboracensis | 1.14 | 8 | 54 |
| Yellow Warbler | Setophaga petechia | 0.97 | 8 | −20 |
| Tropical Kingbird | Tyrannus melanocholicus | 0.84 | 4 | |
| Black-and-white Warbler | Mniotilta varia | 0.76 | 11 | −27 |
| Yellow-throated Vireo | Vireo flavifrons | 0.73 | 9 | 62 |
| Great Crested Flycatcher | Myiarchus crinitus | 0.55 | 8 | 2 |
| Baltimore Oriole | Icterus galbula | 0.41 | 10 | −42 |
| Orchard Oriole | Icterus spurius | 0.10 | 10 | −23 |
We used scores from the Partners-in-Flight [PIF] species assessment and prioritization process that quantifies species-specific vulnerability and conservation need with seven indices reflecting breeding and nonbreeding distributions, relative abundance, threats to breeding, threats to nonbreeding, and population trend (Rosenberg et al., 2016). For each species the seven categories, for which scores ranged from 1 (low priority) to 5 (high priority), were summed to produce a single conservation concern measure between 7 and 35.
All mining concession boundaries were from publicly available sources. Concessions for Peru and Colombia came from Global Forest Watch (2018), who aggregated data from The Instituto Geológico Minero y Metalúrgico and Agencia Nacional de Minería de Colombia for Peru and Colombia, respectively. Ecuadorian mining concessions came from the Ecuadorian Ministry of Mines (Agencia de Regulacion y Control Minero, 2017) and were provided by Daniel C. Thomas of the Rainforest Information Center.
To estimate the global proportion of each species’ abundance within mining concessions, we disaggregated the 8.4km resolution relative abundance estimates to 840m resolution, assuming that birds were evenly distributed across each cell. We then summed the abundance values from cells containing mining concessions and divided by the total abundance summed across all cells. The disaggregation step was necessary to account for the wide range of concessions sizes (roughly 10m2 to 2000km2). In the interest of being conservative in our estimates, we excluded all concessions that were smaller than the 8.4km cells of the abundance data.
We used a Marxan-like approach to identify the minimum set of sites (i.e., least amount of area) that together met a given conservation target, which ranged in 10% increments from protecting 10% of the total abundance of each species to 50% of the total abundance. For all prioritization scenarios, we used the minimum set objective function within the prioritizr R package (Hanson et al., 2018) with the Gurobi optimizer (Gurobi Optimization and LLC, 2018). By prioritizing sites using progressively stricter conservation targets, we were able to rank sites according to their conservation importance. Sites selected at the 10% level are the most important for the protection of these species, while sites only selected at the 50% level, and not lower levels, are the least important.
To determine the relative conservation importance of sites within mining concessions versus the mean importance value of land as a whole in each country, we assigned prioritization importance scores (PIS) of 1, 0.8, 0.6, 0.4, or 0.2 for sites selected at the 10%, 20%, 30%, 40%, or 50% level, respectively. Then, for each country, we calculated the percent difference between the average PIS within concessions (PISmines) and the average score in the country as a whole (PIScountry).
ResultsMining concessions in Colombia, Ecuador, and Peru covered 288,798km2, representing nearly 11% of their combined land area. Ecuador and Peru had the largest percentage of land in concessions, at 15% and 14%, respectively, compared to 6% of Colombia's area.
Concessions disproportionately co-occurred with important non-breeding areas for our focal group of 22 species of Neotropical migratory birds in Colombia and Ecuador, but not Peru (Figs. 1 and 2). Based upon the conservation priority scores of each cell, we determined that Colombian and Ecuadorian mine concessions were located in areas that were 56% (PISmines=0.362, PIScountry=0.231) and 12% (PISmines=0.184, PIScountry=0.165), respectively, more important for conservation than random locations in the country, whereas Peruvian concessions were 74% (PISmines=0.0126, PIScountry=0.0494) less important than random locations for our focal group of birds.

Mines in Ecuador and Colombia, but not Peru, were disproportionately located in areas important for 22 species of Neotropical migratory birds. For each species, the proportion of the avian abundance within mining concessions is scaled by the proportion in the country as a whole. The bias toward important sites is estimated as the difference between this abundance ratio and the proportion of the country's area within mining concessions, with mean bias estimates indicated in red. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of the article.)
 for 22 species of Neotropical migratory birds, based on spatial prioritizations using eBird-generated models, and mining concessions in Colombia, Ecuador, and Peru. Percentages indicate percentage of bird populations that would be supported if the site were protected, with 10% being most important and 50% being least important.")
Importance of sites (i.e. 8.4km×8.4km grid cells) for 22 species of Neotropical migratory birds, based on spatial prioritizations using eBird-generated models, and mining concessions in Colombia, Ecuador, and Peru. Percentages indicate percentage of bird populations that would be supported if the site were protected, with 10% being most important and 50% being least important.
One-quarter of our focal species had >10% of their global populations on concessions during the non-breeding season (Table 1). Species of greater conservation importance and with larger population declines between 1974 and 2014 were most likely to co-occur with mining concessions. For species with >10% of their global nonbreeding populations on concessions, the mean population decline between 1974 and 2014 was 43% (SD=35.6) and species had a mean PIF score of 12 (SD=2), which indicates relatively high concern. This group of species included two that are on both the IUCN Red List and North American Watch List – the “Near Threatened” Olive-sided Flycatcher and the “Vulnerable” Cerulean Warbler – along with the “Watch List” Canada Warbler.
Species with 1–10% of their global populations on mining concessions had mean PIF scores and population declines of 11 (2.4 SD) and 15% (33.5 SD), respectively. The Golden-winged Warbler, which is “Near Threatened” on the Red List and a Watch List species fell in this group. Species with <1% of global populations on mining concessions had mean PIF score of 9 (2.3 SD) and population declines of 8% (37.1 SD).
DiscussionA rapid increase in the extent of mining concessions in the Northern Andes has potentially serious implications for the conservation of biodiversity and ecosystems in the region. We showed that one-quarter of the Neotropical migratory birds examined had >10% of their global populations on land under concession. Moreover, species of high conservation concern and those declining most steeply (mean=43%) – including Olive-sided Flycatcher, Cerulean Warbler, and Canada Warbler – were most likely to co-occur with concessions. The potential loss or degradation of habitat sustaining ≥10% of a global population represents an important benchmark in conservation. For example, habitat sustaining ≥10% of the global population of species ‘Red-listed’ by the International Union for the Conservation of Nature (IUCN) were considered “critical” in a biodiversity guidance note by the International Finance Corporation (2012). Likewise, BirdLife International defines Important Bird Areas globally as sites supporting congregations of ≥1% of the global population of one or more species on a regular or predictable basis.
Although the chance that all concessions transition into active mines may be low, exploration can contribute to habitat degradation via deforestation or degradation, road and camp construction, erosion, sedimentation, disturbance, and pollution (Sonter et al., 2017; Roy et al., 2018). Exploration can also involve clearing areas for the passage of heavy equipment, exposing subsoils, and drilling for sample extraction and analysis (Moon et al., 2005). Abundant evidence demonstrates the potential for dramatic and potentially irreversible effects of habitat loss and fragmentation, road construction, and increased human access on biodiversity and ecosystem function (Forman and Alexander, 1998; Fahrig, 2003). Because mining concessions in two of three countries studied here occurred disproportionately in areas supporting relatively high numbers of migratory birds, exploration and development in them may therefore be expected to have at least some impact on biodiversity conservation in the Northern Andes ecoregion.
One insight that comes from our work is that we must exercise strong caution when generalizing forecasts of impacts across geographies. Even when comparing potential consequences of the same activity across the same region and taxa, we found sharply contrasting patterns of co-occurrence of mine concessions and overwintering areas for birds. Concessions were about an order of magnitude more likely than to overlap important areas for our focal migratory birds in Colombia and Ecuador than in Peru. In our system, the difference may stem from the fact that concessions in Colombia and Ecuador were often in mesic montane forest habitats known to be valuable to Neotropical migratory birds, whereas concessions in Peru were primarily located in arid, landscapes less used by our group of migratory birds. Although our current analysis used small subset of biodiversity features of interest to conservation in the Northern Andes ecoregion, our observation of marked spatial variation in the co-occurrence of mining concessions and habitat supporting Neotropical migratory birds at regional and local scales suggests that more detailed analyses including a wider range of values could best inform the siting and licensing of mine concessions and, eventually, active mines.
Our approach highlights how public data can be effectively integrated to anticipate potential consequences for biodiversity and also to identify locations that are likely to be associated with less severe environmental impacts and/or appropriate for mitigation projects. The prioritization method we used also has the advantage of accounting for complementarity and, as such, facilitates the development of cost- and/or area-efficient portfolios of sites with high conservation value. These planning efforts are not simply intellectual exercises, as a recent survey showed that 74% of prioritizations intended for implementation produced action on-the-ground (Sinclair et al., 2018). Fortunately, recent advances in the spatiotemporal resolution and extent of citizen-science data are transforming the field of systematic prioritizations or spatial conservation planning. Indeed, nearly one-third of downloads of e-Bird data for conservation purposes were used to support conservation plans (Sullivan et al., 2017). In a world struggling to meet the needs of a growing human population amid declining biodiversity and ecosystem health, scientists and practitioners must make use of the new suite of tools that facilitate strategic conservation.
Conflict of interestsNone declared.
ADR and MSM are supported by endowments at the Cornell in front of Lab of Ornithology, RS by a Liber Ero Fellowship, and PA by the Natural Sciences and Engineering Research Council of Canada. We thank J. Mecham for helpful input, A. Spencer for validating our selection of focal species, and D. Thomas for advice and access to data layers with mine concessions. We also are grateful to eBird participants for their data contributions and eBird development team, especially S. Kelling, D. Fink, and T. Auer, for their innovation, support, and helpful discussions. The development of eBird and associated data products and models has been funded by The Leon Levy Foundation, The Wolf Creek Charitable Foundation, NASA (NNH12ZDA001N-ECOF), Microsoft Azure Research Award (CRM: 0518680), and the National Science Foundation (ABI sustaining: DBI-1356308; computing support from CNS-1059284 and CCF-1522054) – all of which were awarded to S. Kelling at the Cornell Lab of Ornithology.

