





As climate and land-use changes threatens biodiversity, the identification of refugia areas for species becomes a crucial strategy in conservation planning. Here, we integrate climate change anomaly with a land-use change model both projected for 2050 to identify refugia areas for 103 bird species that occur in the Brazilian Cerrado. We found that 13% of the Cerrado may serve as refugia for the bird species. In contrast, nearly 35% of the biome might become areas of high risk for those species. Most species (74%) will held from 34% to 85% of their current geographic distribution in areas with less native vegetation, but with low climate anomaly. Apart from the protection of the refugia areas, we suggest restoration of native vegetation in regions that are likely to maintain climate conditions more adequate in the future. These areas should be prioritized to protect places with higher species richness and ease the establishment of corridors that would support climate-induced dispersal from high risk areas to suitable ones. Approaches that allow for the identification and future protection of refugia are fundamental to guarantee the conservation of biodiversity faced with climate change and rapid land-use changes that already taking place.
Climate change is a global threat to biodiversity, being pointed out as the main cause of species extinctions in the next decades (Dawson et al., 2011; Parmesan and Yohe, 2003; Thomas et al., 2004). It is expected that these effects will intensify once climate projections suggest an increase on global temperature of ca. 4.8 °C, depending on the scenario of greenhouse gases emissions till the end of this century (IPCC, 2013). Climatic changes cause variation in the phenology, geographic distribution and composition of ecological communities (Chen et al., 2011; Parmesan, 2006). Land-use change, via conversion, degradation, and fragmentation of habitats was and still is the main driver of biodiversity loss worldwide (Foley et al., 2005; Jetz et al., 2007; Newbold et al., 2015). It is also expected that the combination of climate and land-use change increase even more the extinction rate projected to a near future (Brook et al., 2008; Jetz et al., 2007). For example, habitat loss and fragmentation can reduce the ability of species to change their distribution in pursuit of adequate climatic niches (Brook et al., 2008; Mantyka-Pringle et al., 2012).
Faced with the threats of climate and land-use change, an important strategy to the protection of biodiversity is to identify climatically adequate areas that will maintain suitable habitats for species in the future. One approach to identify these places consists on the use of climate change metrics for the identification of regions that are more or less exposed to these changes throughout time (Beaumont et al., 2011; Garcia et al., 2014; Loarie et al., 2009; Williams et al., 2007). Some examples of climate change metrics commonly used are the climatic anomalies, climatic extremes, and climate change velocity. Climatic anomalies and climatic extremes quantify the magnitude of change in the mean and extreme conditions, respectively, in a determined locality throughout time (Garcia et al., 2014). Climate change velocity is a measure of climatic dislocation rate in the landscape and provides a velocity with which species should move to pursuit their suitable climates (Loarie et al., 2009).
The scenarios of impact of environmental changes on biodiversity in the future focus in climate change and broadly neglects land-use changes (Titeux et al., 2016). Lack of projections that integrate the climate and land-use changes constitutes a great knowledge gap that prevents the development of more trusty scenarios for the implementation of biodiversity conservation policies in the future (Titeux et al., 2017, 2016). Thus, integrating land-use models with climate change metrics allow for the identification of areas that can potentially act as climate and habitat refugia for biodiversity as well as establish more adequate conservation strategies for each area (Alagador and Cerdeira, 2018; Struebig et al., 2015; Triviño et al., 2018; Watson et al., 2013).
The identification and protection of potential refugia in landscapes is an important strategy for conservation planning in the context of global changes (Groves et al., 2012; Morelli et al., 2016; Ribeiro et al., 2018; Stralberg et al., 2018; Struebig et al., 2015). The occurrence of species is strongly affected by climate and their survival depends of the availability of their tolerated habitats, thus, species can face great risk if the climate conditions to which they are adapted to and their tolerated habitats disappear in the future (Garcia et al., 2014; Mantyka-Pringle et al., 2012; Newbold et al., 2015). Tropical species are particularly vulnerable to these changes, as they already live near their maximum thermal tolerance (Araújo et al., 2013; Khaliq et al., 2014), besides having high sensibility and low adaptation capacity (Foden et al., 2013). Owing to high rates of climate and land-use changes expected for the tropics (IPCC, 2013), it is probable that the extent of refugia areas will be the main mechanism by which the species could persist in the future (Reside et al., 2014), as these areas would hold climatic conditions as well as habitats that are more suitable for them.
Here we present a spatially explicit approach which incorporates both climate and land-use change models to identify areas of refugia for bird species in the Brazilian Cerrado. Specifically, our objectives were: (1) to identify major risk areas and possible refugia for bird species in the Cerrado; (2) to quantify the proportion of current geographic distribution of species within each of these areas, and (3) to point out likely conservation strategies according to the level of climate change and land-use in the region.
MethodsStudy area and evaluated speciesThe domain of the Cerrado extends for 23% (200 million acres) of the terrestrial area of Brazil, being the second major biogeographic province of South America and the world’s biggest savannah (Silva and Bates, 2002). It shows a considerable variation in its phytophysiognomy, which includes habitats ranging from open areas to forests of closed canopy (Eiten, 1972). The Cerrado is a Biodiversity Hotspot (Myers et al., 2000), being considered the world’s most threatened savannah (Strassburg et al., 2017). Protected areas cover 8.6% (Vieira et al. 2019) of the Cerrado and only 3% is legally protected by areas of integrated protection (Françoso et al., 2015). Other than the conversion of the major part of native vegetation due to strong farming pressure, the Cerrado finds itself in a predicted future high climate instability region, which makes the biodiversity particularly vulnerable to these changes (Borges et al., 2019; Watson et al., 2013).
The Cerrado shelters 856 bird species (Silva, 1995) of which 30 are endemic (Silva and Bates, 2002) and 56 are threatened according to Brazil’s Red Book on Threatened Species (ICMBio, 2018). Bird species found in the Cerrado show very distinct patterns of geographic distribution, including those of endemic and strict distribution to wide-ranged species (cosmopolitan). Of the 856 species listed for the Cerrado, we included in this study only those species that have the largest proportion of their range within the biome. Species widely distributed in other biomes and having a small proportion of their distribution area within the Cerrado were excluded. Following this criterion, we select a subset of 103 bird species.
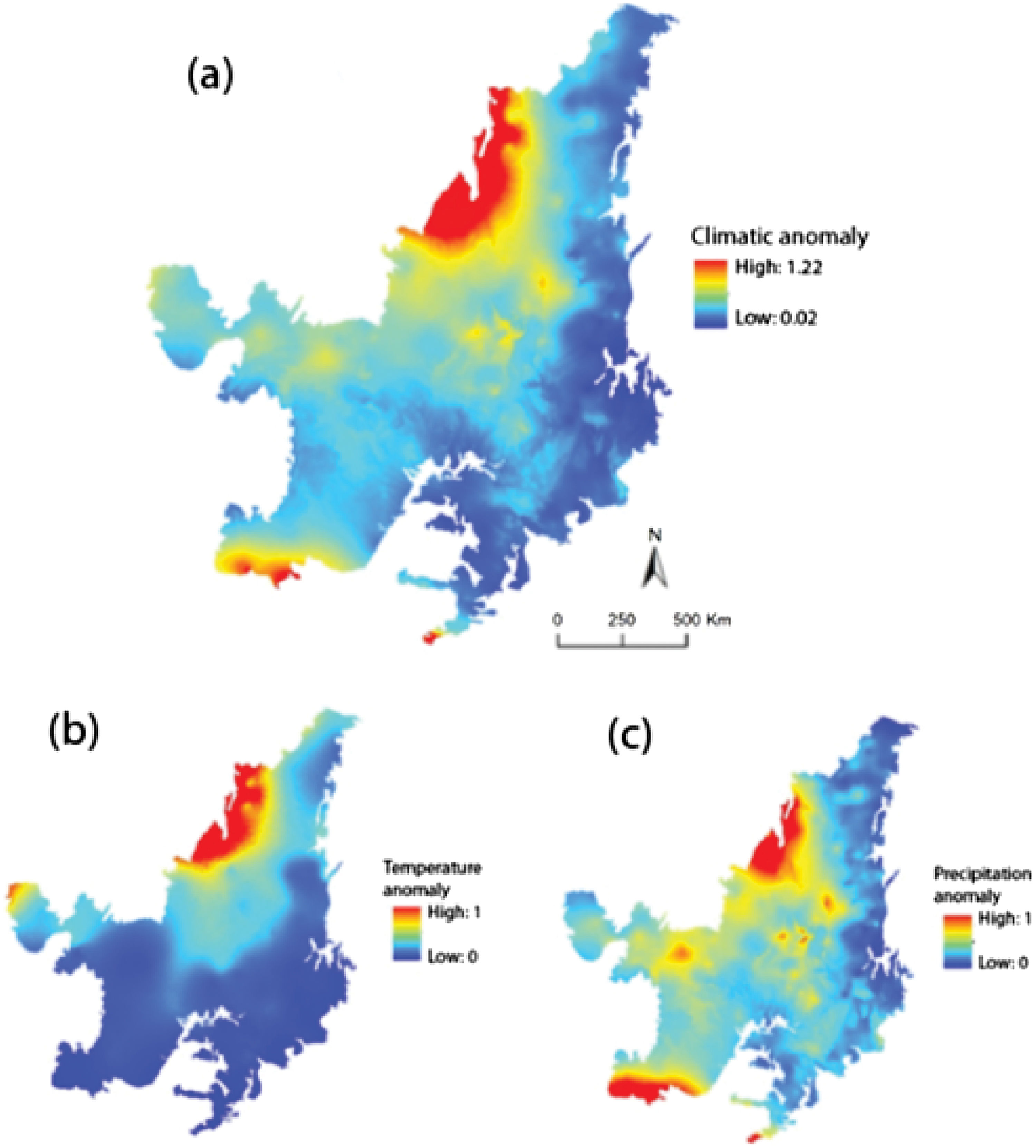
Climate and land-use dataTo quantify the magnitude of local climate change we use standardized local anomalies. We calculated the sum of Standardized Euclidean Distances (SED) for temperature and precipitation between the current period (1960–1990) and 2050 (2041–2060) for each grid cell, according to Williams et al. (2007). Given that standardized local anomalies for temperature and precipitation may show a different spatial pattern, we adopted the same approach as Garcia et al. (2014), who have calculated these values individually. Further, a comparison both sets of results provides a better understanding of which parameter drives the patterns of combined temperature and precipitation change (see Garcia et al., 2014). To evaluate temperature, we use mean annual temperature (Bio1) and for precipitation we use annual precipitation (Bio12). SED differences between the current period and 2050 were standardized by the inter-annual standard deviation (current period) of temperature (seasonality of temperature - Bio4) and precipitation (seasonality of precipitation - Bio15) (Williams et al., 2007). We also calculated the SED for temperature and precipitation individually and standardized all SED values to range from 0 to 1. Higher values of SED indicate higher local climate change (Williams et al., 2007). The four bioclimatic variables mentioned above were obtained in the WorldClim database (Version 1.4; www.worldclim.org/version1) for the present period and 2050 in the resolution of approximately 1 × 1 km (30 s latitude/longitude).
For 2050, we used climate projections of four General Atmosphere-Ocean Circulation Models – AOGCMs: CCSM4, MPI-ESM-LR, HadGEM2-AO and IPSL-CM5A-LR. These AOGCMs were chosen because they belong to groups of models with different predictions and covering the widest range of climatic predictions (Varela et al., 2015). We have chosen to use a scenario of high greenhouse gases emissions (RCP 8.5) compiled by the Coupled Model Intercomparison Project (CMIP) as a standard experimental protocol for studying the output of coupled AOGCMS (available at https://esgf-node.llnl.gov/projects/cmip5/). This seems to be a likely scenario given the trends for greenhouse gas emissions since the year 2000 and, besides, only minor differences have been noticed across all RCPs until 2050 (Diffenbaugh and Field, 2013; IPCC, 2013). To represent the value of each bioclimatic variable in 2050, we use the average of the four AOGCMs.
To evaluate land-use change, we used a map produced by Soares-Filho et al. (2016) projected for 2050. We cropped the map to the extension of the Cerrado and we put it in the same resolution of the climate variables (∼ 1 km x 1 km), since that its original resolution is 500m × 500m. This spatially explicit model simulates the changes in land-use and the carbon emissions associated under diverse scenarios on the demand of agricultural land and deforestation policies for Brazil (for more details see Soares-Filho et al., 2016). We classified grid cells with different uses of land into two categories: “with more native vegetation” and “with less native vegetation”. Cells occupied by savannas, savannas in PAs, forests and forests in PAs were classified as having more nature vegetation (land use refugia), whereas cells occupied with other land uses (urban and agricultural) were classified as transformed (less native) vegetation.
Land-use and climate change integrated riskTo integrate the climate anomaly map with the land-use change map, we divided the climatic anomaly values into two categories: values above the median (high climatic anomaly) and values below the median (low climatic anomaly, i.e. climate refugia). Then, we overlapped the two maps to identify the regions with different combinations of climate and land-use classes (Fig. 1). In this paper, we define refugia as those areas that might have a combination of low projected climatic anomaly and also projected retention of native vegetation until 2050. This is a wider understanding of refugia compared to its original definition, but justifiable as a way to include land-use changes on it.
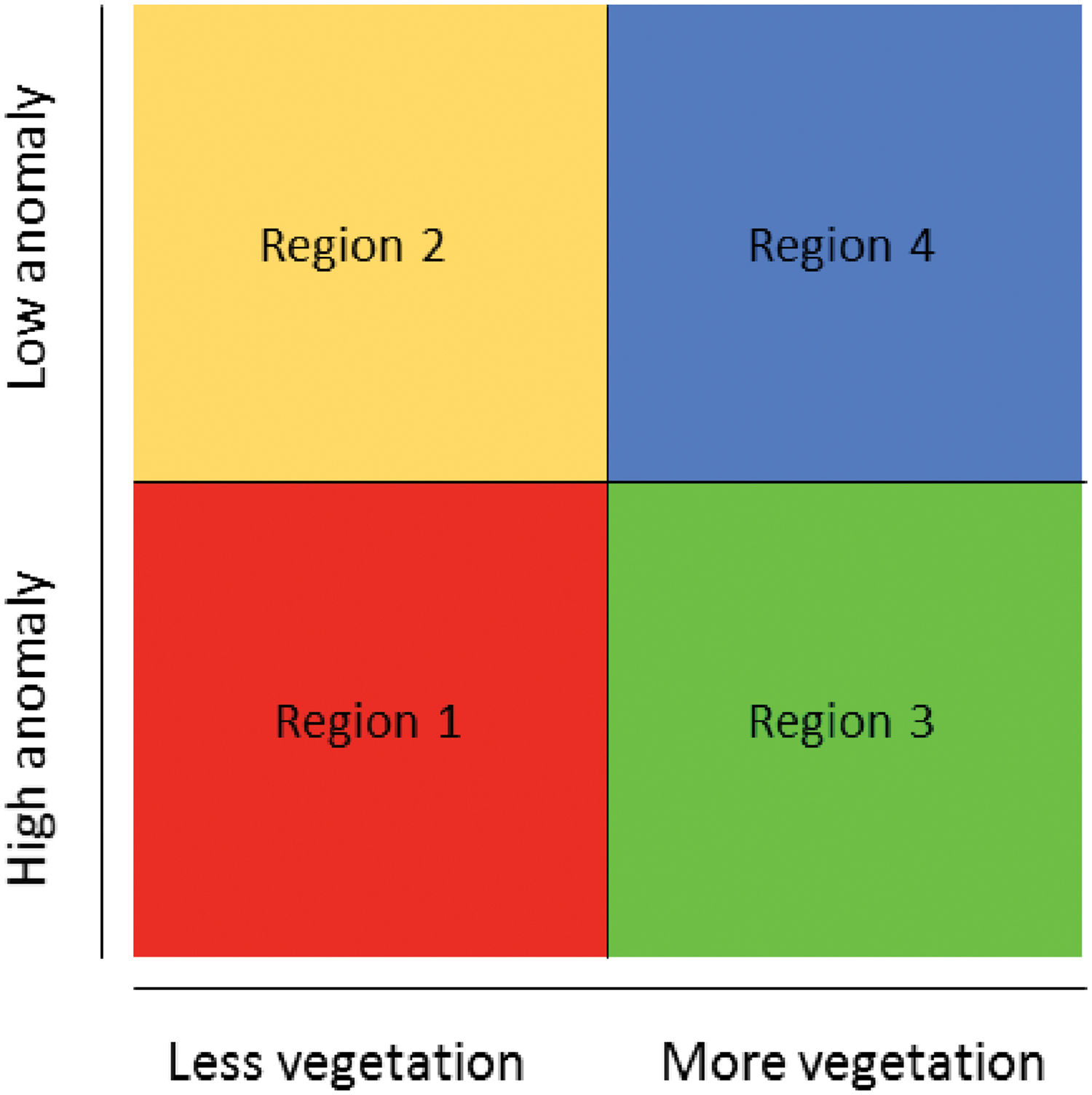
Conceptual diagram showing the intersection between the climate anomaly and land-use projected for the Cerrado until 2050. Region 1: places with less native vegetation and high climatic anomaly – high risk; Region 2: places with less native vegetation and low climatic anomaly; Region 3: places with more native vegetation and high climatic anomaly; Region 4: places with more native vegetation and low climatic anomaly – refugia.
We calculated the proportion of geographic distribution of each species and those listed as threatened by the national list that occurs inside each of the four regions presented in Fig. 1. For that, we used the species range maps made available by BirdLife International (available at http://www.birdlife.org/datazone/info/spcdownload). Then, we rasterized species range maps and put them on the same resolution as our map which combines climate and land-use changes. We calculated the proportion of species distribution area that occur inside each of the regions and also calculated how much of refugia areas occur inside protected areas currently established in the Cerrado. Hereafter we refer to refugia as areas considered as both climate and land use refugia.
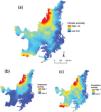
ResultsCerrado regions that were possibly exposed to higher values of climate anomaly (temperature + precipitation) would be localized in the north and northwest, and in smaller proportion in the south and southwest (Fig. 2a). Looking at patterns of temperature and precipitation changes separately; we observed an overlap of both regions in the north and northwest (Fig. 2b and c). A small portion in the west region would also be exposed only to the major values of temperature anomaly (Fig. 2b), while some patches in the center and portions of the southwest and south regions would be exposed only to the highest values of precipitation anomaly (Fig. 2c). Our land-use model projected for 2050 shows that the north region would concentrate the largest part of native vegetation, while south and west regions would be the ones losing more native vegetation (Supplementary material, Figure S1).
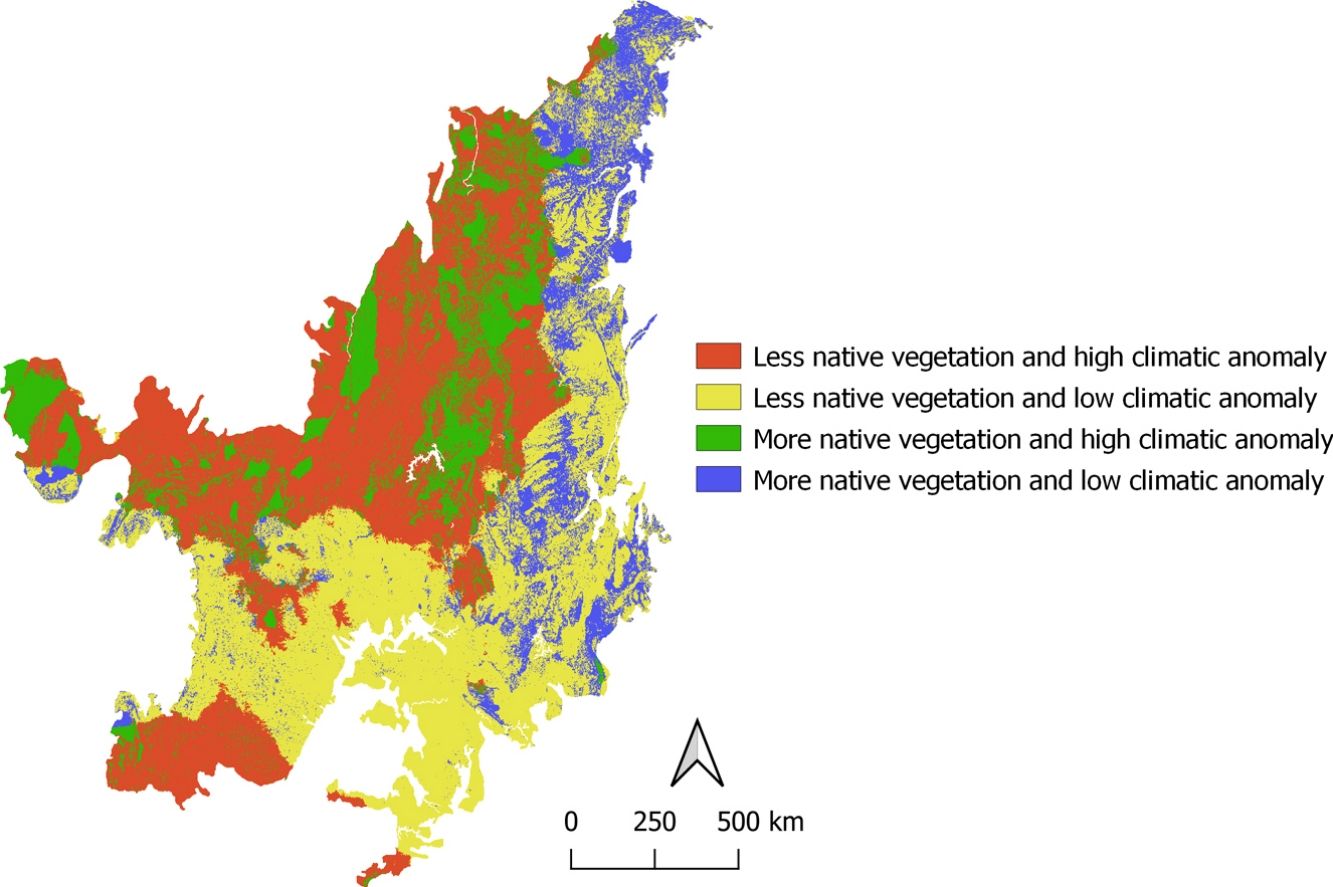
When we combined data from climatic anomaly and land-use changes, we found that 35.4% (∼71 million hectares) of the Cerrado would be exposed to high climatic anomalies and poor native vegetation until 2050 (i.e. high-risk areas). High-risk areas are found in the north, northwest, middle, west and small areas in the south and southwest regions (Fig. 3a).
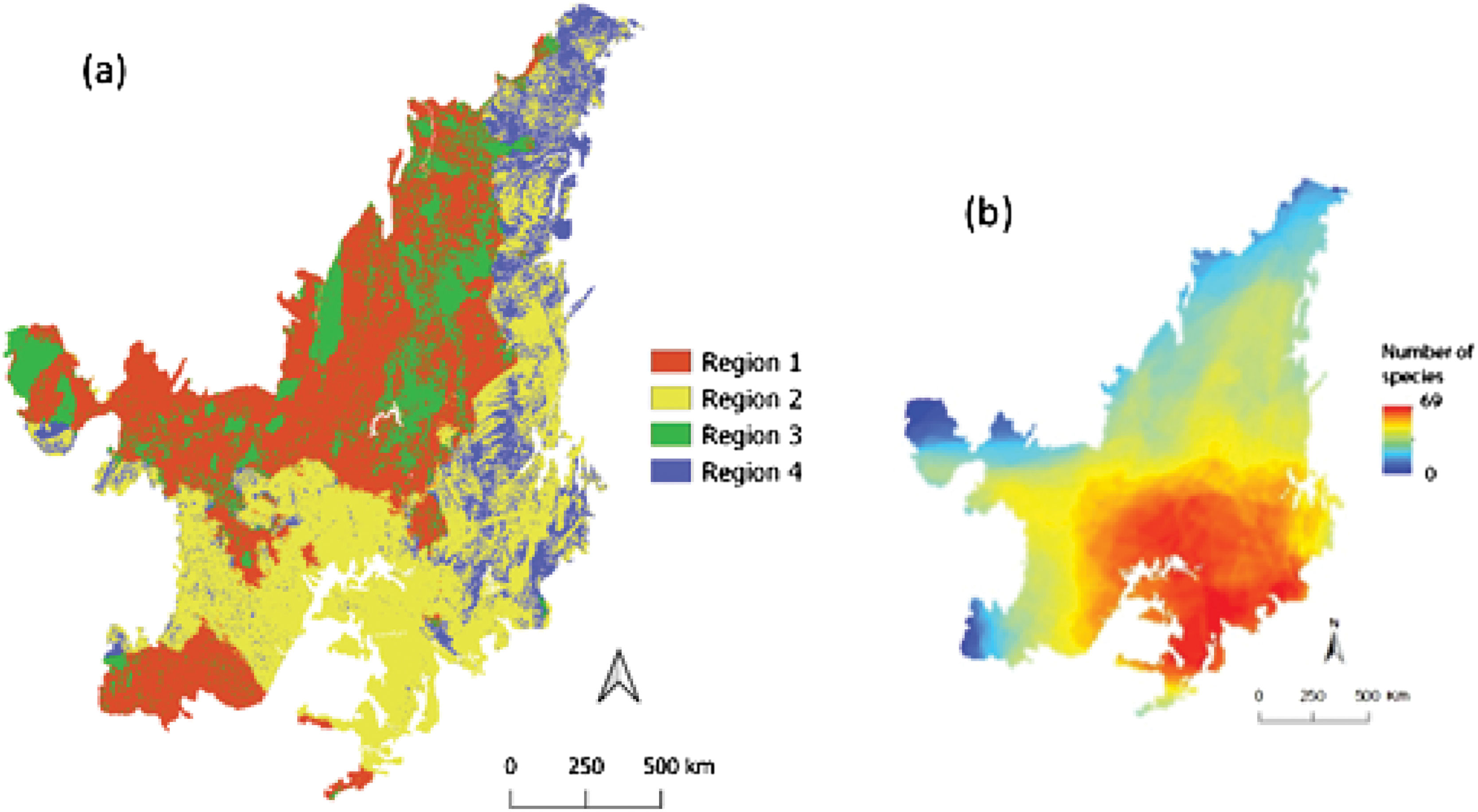
Refugia and high-risk areas according to the climate and land-use changes projected for the Cerrado until 2050 (a) and present richness of bird species (b). Region 1: places with less native vegetation and high climatic anomaly – high risk; Region 2: places with less native vegetation and low climatic anomaly; Region 3: places with more native vegetation and high climatic anomaly; Region 4: places with more native vegetation and low climatic anomaly – refugia.
Areas likely to retain their native vegetation with low climatic anomaly (i.e. potential refugia), cover only 13% of the Cerrado. These areas are in the east part, from north to the south of the Cerrado (Fig. 3a). Areas with less native vegetation and with low climatic anomaly (37.5%) will be in the south between southeast and southwest, while areas holding much native vegetation and high climatic anomalies (14%) will be in the middle between the north and western regions (Fig. 3a).
Nearly 74% of bird species will occur inside areas with less native vegetation, but with low climatic anomaly until 2050 (from 34% to 85% of their ranges; see Supplementary material, Table S1). Twenty-one species (20.4%) will keep from 30% to 69% of their distribution in areas with less native vegetation and high climatic anomaly. We found that only four species (Celeus ochraceus, Asthenes luizae, Augastes scutatus and Embernagra longicauda) will have most of their distribution (49% to 64%) in areas with native vegetation and low climatic anomaly (i.e. potential refugia) (Table S1). Almost 100% of the species (n = 94) will have a largest part of their distribution areas inside regions with less native vegetation, while 22.3% of the species will have a large part of their distribution areas inside the regions with high climate anomaly (Table S1).
Areas of high species richness (Fig. 3b) will fall outside future refugia. On average, the evaluated species will have 27% of their distribution in Region 1, 46% in Region 2, 12% in Region 3 and 15% in Region 4 (Fig. 4a).
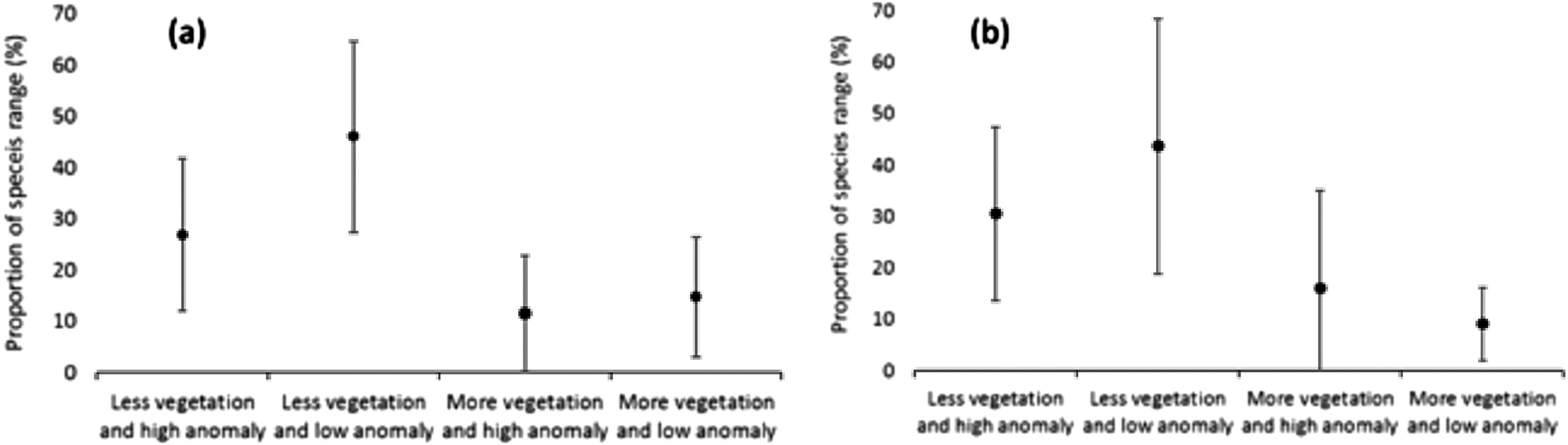
Among studied species, 15 are considered threatened by the national list of threatened species. These species follow the same pattern of the total set, in which, on average, will have 31% of their distribution areas in Region 1, 44% in Region 2, 16% in Region 3 and only 9% in Region 4 (Fig. 4b). Four species (Cercomacra ferdinandi, Paroaria baeri, Pyrrhura pfrimeri and Synallaxis simoni) will be totally out of the refugia areas, being that the four are endemic and two of them (C. ferdinandi and P. pfrimeri) threatened (Table S1). Only 11.4% of the refugia areas occur inside protected areas.
DiscussionWe combined projection of climate anomaly with a model of land-use change and identified refugia areas that safeguard bird species from negative impacts of environmental change over the next decades. We also showed that most species would be vulnerable as only a small part of their distribution would lie within protected refugia areas. Here we discuss these results and likely conservation strategies for regions with different combinations of climate and land-use change.
Identification of future biodiversity refugia areas is a fundamental strategy used by managers and conservation practitioners faced with the threat of environmental alterations. However, most studies based on this approach focus only on climate change scenarios, neglecting land use changes (Titeux et al., 2016). As a consequence, decision makers end up having access to less trustworthy scenarios, because when land-use change is included in the analyses, predicted suitable areas for species change and/or reduce significantly (Struebig et al., 2015; Triviño et al., 2018).
Our approach showed that only 13% of the Cerrado will preserve habitat and stable climate conditions, being able to act as refugia for the species until 2050. Moreover, most species have large portions their distributions in high risk areas. Bird populations in these areas can be impacted in many ways: land-use change can increase population decline (Cavalcanti, 1999; Lopes et al., 2010) and increase predation and nest parasitism (Borges and Marini, 2010), local climatic changes can induce change in species’ distribution, physiology and behavior, as well as alter the periods for activities such as migration and reproduction, favor the emergence of new diseases and biotic interactions, with negative consequences for the demography and dynamics of population (Crick, 2004; Sekercioglu et al., 2012). Further, it is known that species that inhabit tropical regions are particularly more vulnerable to climate change, once that they have already experienced near maximum tolerance levels of their temperature conditions (Araújo et al., 2013; Khaliq et al., 2014), have low capacity of physiological adaptation and plasticity (Araújo et al., 2013; Hoffmann et al., 2013). Due to differences in their biological and ecological traits, some Cerrado bird species would be more vulnerable to climate and land use changes than others (Borges et al., 2019).
We found that four species (C. ferdinandi, P. baeri, P. pfrimeri and S. simoni) will be the most vulnerable, as they occur totally outside refugia. These species should be prioritized for conservation and have their population monitored, especially because they are endemic to the Cerrado. C. ferdinandi, and P. pfrimeri are also threatened with extinction and were classified as highly vulnerable due to their high exposure and low responsiveness to climate and land use changes (Borges et al., 2019). C. ferdinandi and S. simoni will occur mainly in regions with more native vegetation and high climatic anomaly (Region 3, in Fig. 4), while P. baeri and P. pfrimeri have larger distribution in the region with high risk, i.e. less native vegetation and high climatic anomaly (Region 1, on Fig. 4). Because they occur in different regions, protecting these species demand different conservation strategies (Watson et al., 2013). With exception of P. pfrimeri that occurs in a thin band of dry forest, near Serra Geral, the other three species have overlapped distribution in the Araguaia River’s hydrographic basin (del Hoyo, et al., 2018). The preservation of these areas is critical, especially because habitat loss is the main threat to the populations of these species in the Cerrado (del Hoyo et al., 2018).
Different conservation strategies should be developed in regions likely to present different combinations of climate and land-use change (see Watson et al., 2013). For example, in areas with less native vegetation and high climatic anomaly, there is need for monitoring populations to identify current threats and identify the most vulnerable species that would demand more assisted conservation interventions, like future translocations to refugia areas. In areas with low suitability that pose risks to species permanence, management of the surrounding landscape to establish corridors and steppingstones which facilitate the movement and dispersal of species in search of suitable conditions is a priority action (Hole et al., 2011).
In areas with less native vegetation but low climatic anomaly, restoration of those areas and their neighboring ones is important as a strategy to increase connectivity and increase the size of populations by facilitating species dispersal. In areas where native vegetation is abundant but might face high climatic anomaly, it would be important to reduce the existing threats and guarantee the preservation of that native vegetation so that the species have opportunities to adapt to local climate or disperse to refugia areas with more adequate climate.
Finally, in areas with more native vegetation and low climatic anomaly, the creation of new protected areas and avoidance of current threats such as deforestation, wildfires and species invasion would guarantee the permanence of bird species in a shorter and a longer time period (Watson et al., 2013). These areas of refugia may require conflicting management actions, as they need to maintain viable populations of resident species, while at the same time, providing conditions that facilitate the settlement of new colonizers from unsuitable areas (Hole et al., 2011). The possibility of establishment of new communities with new sets of species need to be previously assessed with caution as, depending on species, it is likely to have significant impacts on local equilibria (Jackson and Sax, 2010). In order to encourage the protection of potential refugia areas, it is important to financially support landowners in preserving remnants of native vegetation through payment for environmental services, tax relief or other compensatory measures that are of interest to them.
Species-rich areas in the Cerrado overlap areas projected to lose native vegetation, but with low climate anomaly in the future (especially in southern Cerrado). Therefore, restoration becomes a fundamental strategy for the protection of Cerrado bird species. Currently 26% of the biome found in private lands must be restored to comply with the Brazilian law on the protection of native vegetation (Vieira et al., 2018). It is already possible to develop a sustainable future for the Cerrado through the increase in productivity of cattle raising in the region and restoration of degraded pasturelands (Strassburg et al., 2017). This would save all of the necessary land for agricultural expansion, increase the production of bovine meat in 49% and still open 6.38 million hectares for restoration (Strassburg et al., 2017). The restoration of these areas is primordial to avoid not only the loss of the Cerrado’s biodiversity, but also the loss of ecosystem services (Vieira et al., 2018). For areas in region 2 of our scheme (areas with less native vegetation and low climatic anomaly), it would be important to provide financial and fiscal incentives to landowners – especially smallholders – to support their compliance to the native vegetation law in Brazil. This compliance would be achieved mainly through restoration, and once these regions would have adequate climate conditions, restoring these regions would ensure the occurrence of resident species and also migrants arriving from regions of less suitable or even unsuitable climates.
Given the low resolution of the climatic models at our scale of analysis, it is not possible to evaluate if our refugia areas have local characteristics (e.g. streams, lakes, cold air drains and topographic exposure to radiation and wind) which could create microrefugia in an even thinner scale that could favor the species survival (Ashcroft, 2010; Gavin et al., 2014). However, for the species that present occurrences in more sparse areas such as birds, it is expected that the bigger scale refugia (macrorefugia) provide a better and sparser protection throughout time (Ashcroft, 2010). A caveat of our study is the spatially coarse definition of refugia, so that some specific sites within the defined refugia region may present important changes that some species will not be able to tolerate. However, the conservation strategies outlined for each of the four regions identified in our study are broad and generic suggestions rather than detailed and mandatory actions for a specific location (for an analysis considering the particular requirements for each species see, (Alagador et al., 2016). Approaches that are able to identify areas that are most suitable for species, considering the future threats of climate and land use changes, as presented here (see also Struebig et al., 2015; Triviño et al., 2018), remain critical to biodiversity conservation planning.
Conflict of interestNone.
We thank Bruno Ribeiro for helping with the figures. RL research is funded by CNPq (grant #306694/2018-2). FJAB received a PhD scholarship from CNPq. This paper is a contribution of the INCT in Ecology, Evolution and Biodiversity Conservation founded by MCTIC/CNPq/FAPEG (grant 465610/2014-5). Three anonymous reviewers provided comments that greatly improved the quality of this manuscript.

