







Fire is a global disturbance that has a strong influence on biodiversity. Seasonal fires are common in neotropical savannas of South America, yet few studies have analyzed how the fauna of these habitats and nearby forests respond to fire. We evaluated the effects of fire on the community of non-volant small mammals in the Colombian llanos by comparing their populations in both unburned (no record of fire for the last 20 years) and recently burned areas (three years after the last fire event). Vegetation structure was monitored at 240 plots. We used hierarchical single-season occupancy modeling to determine potential effects of fire on small mammal detection and occupancy. Our results indicate that fire has long-term effects on plant diversity and richness, as well as on the species composition. Three years after the last fire event, the average richness of plant species at burned sites was lower than at unburned sites. Fire also affected the distribution of vegetation strata leading to an increased abundance and diversity of plants in the ground and herbaceous strata in burned plots. The hierarchical models indicate that mammals’ occupancy tracks these changes in vegetation structure. However, the effect of fire was not consistent among species. We found that fire may restrict (Didelphis marsupialis), promote (Zygodontomys brevicauda) or have no impact (Oecomys sp.) on the mammals’ distribution depending on how each species uses the strata that disappeared because of fire. These findings provide evidence toward wildfire prevention for biodiversity conservation in fire-sensitive ecosystems of northern South America.
Fire is an agent of disturbance in almost all the biomes of the world, and it has a pivotal role in terrestrial and atmospheric systems (Bowman et al., 2013; Jolly et al., 2015; Pausas and Keeley, 2009). Fire can contribute to the functioning of many ecosystems and the maintenance of biodiversity (Boesing et al., 2017; Kelly et al., 2016, 2012) by influencing the spatio-temporal patterns of landscapes (Chia et al., 2015), the structure of vegetation (Oliveras and Malhi, 2016; Pausas and Ribeiro, 2013), and the distribution and abundance of fauna (Briani et al., 2004; Litt and Steidl, 2011; Mowat et al., 2015).
Climate change and anthropogenic activities are increasing the frequency and number of fires worldwide (Aragão et al., 2018; Armenteras and Retana, 2012; Pausas and Keeley, 2009). In tropical regions, the intervals between fire events are becoming shorter while wildfires are getting more intense (Bowman et al., 2013). Such shifts in fire regimes are deeply affecting vegetation structure (Armenteras et al., 2021; Barlow et al., 2002; Barlow and Peres, 2008) by altering the natural successional trajectories and by favoring the arrival of understory herbs and grasses that, amongst others, increase forest flammability (Armenteras et al., 2021; Mowat et al., 2015). Fire also acts as an agent of disturbance for animal communities (Pausas, 2019) both indirectly, by altering the structure and composition of vegetation (Hohnen et al., 2015; Litt and Steidl, 2011; Mowat et al., 2015), or directly by causing death by extreme heat, smoke or direct contact (Peres, 1999). Nevertheless, our understanding of how fauna responds to fire disturbances is still limited in the tropics, compared to our knowledge on the responses of vegetation (de Andrade et al., 2014; Pausas, 2019; Pausas and Parr, 2018) but fire is known to exert great impacts over the distribution, diversity, and abundance of many species (Roberts et al., 2011; Chia et al., 2016).
Faunal responses to fire are diverse, for they are strongly related to their own life-history traits. In the short-term, for instance, the primary response is to escape from fire by moving to unburned sites or by using refuges (Kelly et al., 2012; Pausas, 2019; Pausas and Parr, 2018). Traits such as small body size or high dispersal ability define these species. However, responses, especially in the mid and long-term, are less clear and little is known about the life-history traits that allow species to persist or recolonize after fire disturbances (Driscoll et al., 2010; Sutherland and Dickman, 1999). Particularly, there is a lack of knowledge of the effects of fire regimes on biodiversity (Lindenmayer et al., 2016), on the responses of fauna at large spatial scales (Brotons et al., 2008), and how fire-induced spatial heterogeneity affects animal distributions, occupancy and their persistence (Chia et al., 2015; Driscoll et al., 2010; Lindenmayer et al., 2013).
In the neotropical region, previous studies showed how forest fires affect the post-fire successional patterns of small mammals in the Cerrado ecoregion, where different species appear at specific post-fire stages (Briani et al., 2004; Henriques et al., 2006). Other studies have analyzed the use of habitat by rodents before and after a fire (Vieira and Marinho-Filho, 1998), as well as the relationship of small mammal communities with different regimes of fire (Vieira, 1999) and the immediate post-fire changes of rodent abundance and diet (Vieira and Briani, 2013). More recent studies in the Cerrado ecoregion have found that the loss of canopy cover because of fire favors the invasion of generalist species at the expense of specialist species (Camargo et al., 2018). All these findings suggest that there is a closed relationship between vegetation structure and species occurrence during the time that follows a fire event.
Studies that analyze fire impacts on fauna have focused on target groups such as mammals and birds (Bellard et al., 2012). The non-volant small mammals (i.e. rodents and marsupials) have important functional roles in several ecosystems, acting as common prey of other mammals, birds, and reptiles (Cheveau et al., 2004; Kotler et al., 1993), as seed dispersers and predators (Jansen et al., 2002; Muñoz and Bonal, 2011), and influencing soil aeration, etc. (Campos et al., 2017; Litt and Steidl, 2011). Although this group has large taxonomical and functional information gaps in the neotropical region (Gardner et al., 2008; Voss, 1991), the non-volant small mammals may represent an ideal subset for evaluating the short, medium and long-term responses of fauna to fire disturbances since their occurrence depends on vegetation structure and their habitat and food requirements can be severely affected by fire (Banks et al., 2011; Roberts et al., 2015). For example, they use the structural elements provided by vegetation at a microhabitat scale for recolonizing affected areas (Plavsic, 2014), for habitat use (Brotons et al., 2008; Chia et al., 2015; Fox, 1982), for feeding, and for refuge (Monamy and Fox, 2000), as well as for exploiting temporary resources (Brotons et al., 2008). Hence, several responses might be expected depending on species life histories: for example, it is expected that the abundance and richness of species adapted to dense vegetation and arboreal strata decline in burned areas, as higher strata are the first to disappear because of fire. Meanwhile, the abundance and richness of species adapted to open habitats are expected to increase as the lower strata persist after a fire event (Chia et al., 2016; Litt and Steidl, 2011).
Here, we assessed the mid-term effects of fire on non-volant small mammal communities of the Colombian Orinoco region. This region is one of the ecologically most threatened regions in northern South America because of climate change, land management practices, and the development of often unsustainable economic activities, (i.e. live-stock ranching, crops, and oil exploitation, among others) (Romero-Ruiz et al., 2012), that use fire as a tool for clearing land (Armenteras et al., 2005). This increases the risk of uncontrolled wildfires, alters fire regimes, and degrades natural savannas and adjacent forests areas (Armenteras et al., 2021; Romero-Ruiz et al., 2010). We hypothesized that species composition, abundance, and detectability will change depending on species responses to fire and vegetation structure. We also predicted that unburned forest would show higher occupancy probabilities than burned forest.
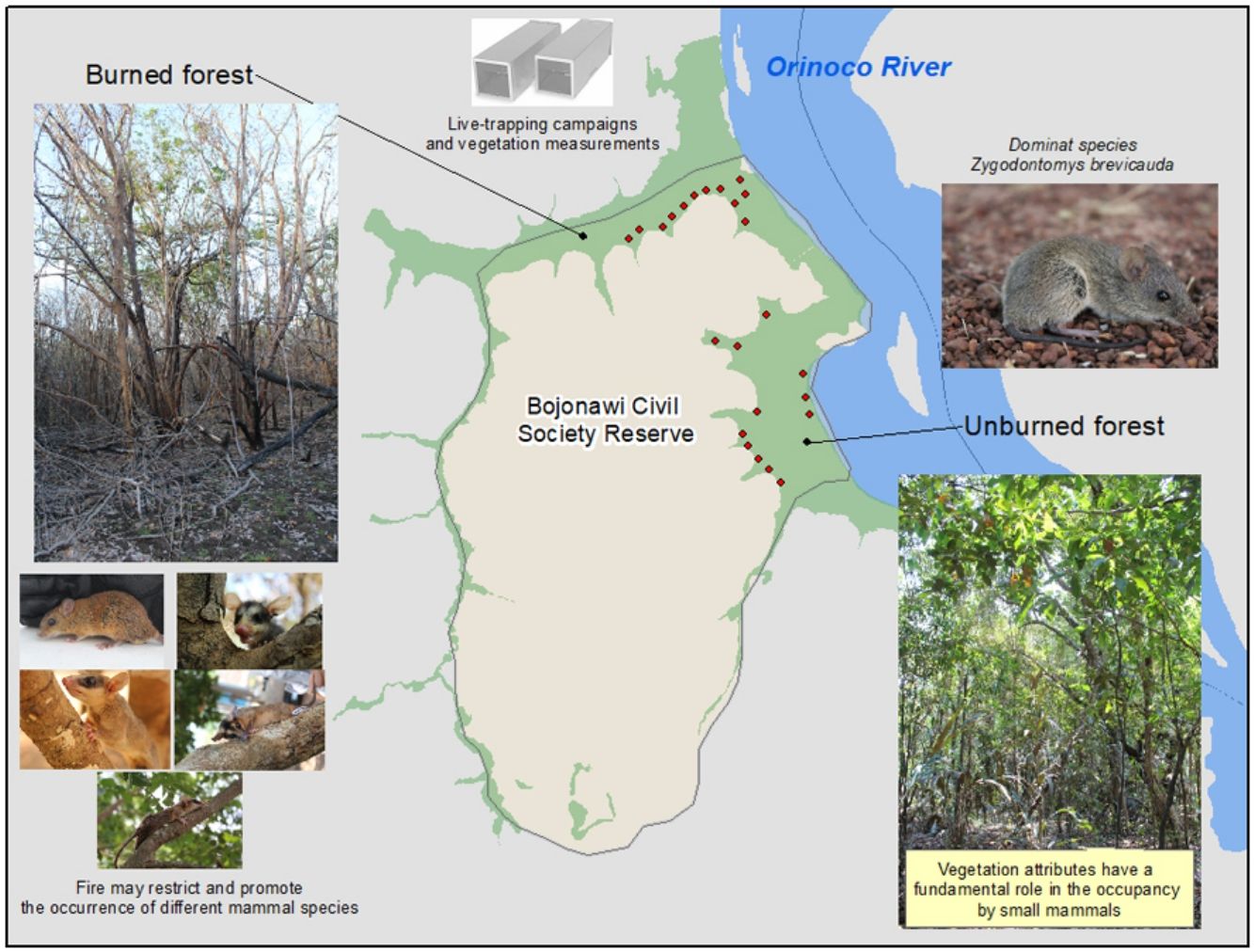
MethodsStudy siteThe Orinoco basin has a vast richness of fauna and flora, providing multiple ecosystem services such as below-ground carbon storage, feeding resources for domestic and wild animals and habitat for charismatic species (Buisson et al., 2019). The Orinoco region comprises at least 70 species of small mammals including 26 species of marsupials (Ferrer Pérez et al., 2009) that inhabit fire adapted and fire sensible ecosystems such as neotropical savannas and gallery forests, respectively (Armenteras et al., 2021). We conducted this study in the Bojonawi Civil Society Reserve (Fig. 1), a protected area that is located in the second largest savanna system in South America after Brazil (Hernández and Sánchez, 1992; Romero-Ruiz et al., 2012).

The Bojonawi reserve belongs to the Colombian Orinoco llanos, municipality of Puerto Carreño (6°07′ and 6°02′N 67°29′ and 67°34′W), and it encompasses an area of 4680ha (Trujillo et al., 2008). It has a tropical climate, with a uniform temperature throughout the year and a monomodal precipitation regime ranging from 1000mm to 3500mm/year (Correa et al., 2005). The dry season takes place between December and April, during which most of the fires occur; and the wet season occurs between May and November, when gallery forests and most of the savannas are flooded. In 2015, close to 80% of the Reserve was burned as a consequence of an accidental wildfire during the dry season, affecting most of the savannas, and approximately 350ha of the forest.
SamplingThe sampling of non-volant small mammals was carried out three years after the last fire in the study area. The survey period covered the beginning (November of 2018 and 2019) and end (March–April of 2019 and 2020) of the dry period and included a total of 24 sampling sites randomly distributed across the Bojonawi reserve (Fig. 1). A half of the sites (12) were located at unburned sections (UB) and the other half at burned sections (B). In each treatment (unburned-burned), six sampling sites were located between 80 and 100m from the forest edge, and six sampling sites at the forest's interior (Fig. 2). We chose this arrangement in two different areas (edge-interior) of the gallery forests to encompass the extent of variability in the vegetation structure. Unburned sites were considered as controls since these areas remain unburned for at least the last 20 years. Fire history was reconstructed using satellite imagery, official vegetation cartography, and data on thermal abnormalities seven years before sampling (2010–2017). Data was gathered from the Fire Information for Resource Management System (FIRMS) as well as from MODIS and VIRS sensors and previous vegetation studies (Armenteras et al., 2021).
. L means large size Sherman traps (10×12×38cm), M means a medium size Sherman trap (8×9×30cm), and S means small size Sherman trap (8×9×23cm). Real photographs of unburned – UB (b) and burned areas – B (c).")
Sampling scheme in the savanna gallery forest landscape of the Bojowani reserve, Colombia (a). L means large size Sherman traps (10×12×38cm), M means a medium size Sherman trap (8×9×30cm), and S means small size Sherman trap (8×9×23cm). Real photographs of unburned – UB (b) and burned areas – B (c).
Although sampling sites had the same general physiognomy and similar climatic conditions, they can be considered as independent sampling units since they were separated by a considerable distance. The maximum and mean distance between contiguous sampling sites was 1000 and 304m, respectively. Such a distance between contiguous sampling sites is not easily covered by South American Cratidae species, whose home ranges (ranging from 70 to 1000m2, Alho et al., 1986; Giuggioli et al., 2005) and maximum daily movement (around 100m, Maroli et al., 2020) tend to be smaller.
Sampling sites consisted of a set of 10 Sherman live-traps (5 traps of 8×9×23cm, 4 traps of 10×12×38cm, and 1 trap of 8×9×30cm; H.B. Sherman Traps Inc., Tallahassee, Florida, USA). At each sampling site, traps were deployed on the ground following two parallel lines encompassing a total area of 900m2; each line consisted of 5 traps spaced apart by 15m and each line was spaced apart by 15m as well. Each trap was georeferenced, and its location was marked with a green plastic tube that allowed us to place the traps at the same spot during each trapping period. Sherman traps were baited with a mix of peanut butter, oats, sardines, and banana essence. At burned locations, the traps were covered with litter to avoid detection by potential predators and overheating of the captured animals.
At each sampling campaign, the traps were activated at dusk, checked daily at dawn, and rebaited during the day. Each trapping campaign lasted for 11 continuous nights. The total capture effort was 2640 trap nights per trapping period (10,560 traps nights in total).
For each captured individual we registered the date, trap location, genus (or species when possible), weight, sex, age (juvenile or adult) and reproductive stage (inactive, pregnant, inguinal testis or scrotum testis) (Hoffmann et al., 2010). The fur of each captured individual was marked with a unique clipped hair pattern to identify recaptures within each sampling period and then was liberated at the exact place where it was captured. Field identification was carried out following the protocols for morphological characters and distributions established by Emmons and Feer (1997); Cuartas-Calle and Muñoz-Arango (2003); Gardner (2008) and Patton et al. (2015) for most of the captured rodents and marsupials, and Voss (1991) for the rodents that make up the Zygodontomys genus.
Those individuals that could not be identified in the field were sacrificed using an injection of lidocaine in the caudal artery and collected for further identification in the laboratory. These specimens were preserved by preparing their skin and skull that were then deposited in the mammal collection “Alberto Cadena Garcia” from the Natural Science Institute (ICN) of the National University of Colombia – Bogotá. Identification followed protocols suggested for the external and cranial morphological characters in Patton et al. (2015) for South America rodents and by Gardner (2008) for South America marsupials. Specifically, we followed Weksler (2015) to determine the genus from the Oryzomyini tribe. Carleton and Musser (2015) was used to determine species of Oecomys, and Voss (1991, 2015) was used to determine the species of Zygodontomis. We confirmed the specimen's identification by comparing our collected specimens with the deposited material in the mammal collection of the ICN and by consulting with experts.
This study was supported by the research permit No. 0255 of March 14, 2014 of the National Authority of Environmental Licenses (ANLA) and endorsed by the Ethics committee of the Sciences Faculty of the National University of Colombia – Bogotá, certificate 02-2018 and 07-2019. All the field and laboratory methods followed the guidelines established by the American Society of Mammologists for the use of wild mammals for research and education (Sikes, 2016).
Vegetation structureTo describe the vegetation structure, we estimated plant species diversity and structural diversity at each sampling site. A circular plot of 1m2 was established using each Sherman trap as the center point (Sunyer et al., 2015), so that the vegetation was estimated at a fine-scale in all 240 plots. Moreover, the collected information at the trap scale was averaged from the 10 traps of each sampling site, covering an area of 900m2, to have a measure of vegetation structure at the sampling site level. We followed Rangel and Lozano (1986) to classify vegetation strata according to its height: ground <0.3m, herbaceous (h) range from 0.3 to 1.5m, shrub (ar) from 1.5 to 5m, subarboreal (Ar) from 5 to 12m, low arboreal (Ai) from 12 to 25m, and high arboreal >25m. The cover percentage of the woody debris was also assessed. At every site, we identified and counted the number of plant species that may provide food for small mammals (seed, fruit, leaf, sprout), with support of local botanists and ecologists. Although this measure does not provide an accurate estimation on the actual food resources, given the diversity of small mammals, the different phenology in fruit production, etc., it may represent a proxy of potential food availability for the community of small mammals across the study area.
Data analysisPlant species diversity was estimated by calculating the first three Hill numbers (q=0,1,2). The Hill numbers represent a mathematically unified family of diversity indices that incorporate the variability in relative abundance and species richness by changing the exponent q (Jost, 2007, 2006). When q=0, the species abundances do not count at all and the species richness is obtained. When q=1, the species are weighed in proportion to their frequencies, and the index value can be interpreted as the effective number of common species in the community. When q=2, abundant species are favored, and rare species are discounted. Thus, index values are equivalent to the inverse Simpson concentration. Differences in plant diversity between burned and unburned sites were tested using t-tests. The structural diversity of vegetation was estimated by means of a principal component analysis (PCA) using information about the coverages (%) of vegetation strata. We retained the first two components (pca1 and pca2) as covariates of hierarchical models.
To determine the mid-term effects of fire, we assessed the changes in the composition, abundance, and detectability of small mammal communities in burned and unburned gallery forests four years after fire. To account for low capture rates (several zeros in datasets), we employed hierarchical occupancy models (MacKenzie et al., 2002). These models allow for more precise estimates of occupancy by accounting for differences in species detectability (i.e., a great number of absences) at different scales. These models have been used for addressing ecological questions such as species presence in response to fire disturbances, providing probabilistic information of the species presence in relationship to predictable variables (Anderson et al., 2016; Kéry and Royle, 2008; MacKenzie et al., 2002).
We did not include recaptures in statistical models because of the low-recapture rates. Hierarchical models consist of one submodel to describe the ecological process (or ‘state’) and another submodel for the observational process, which is conditional by the ecological process (Kéry and Royle, 2016). The ecological process is modeled as a Bernoulli random variable governed by the ‘success probability’ Ψ:
where zi is a latent variable representing the true state of occurrence at site i (i=1,…,24 in this study) and the Bernoulli parameter Ψ is the probability of occupancy or the expected value of z.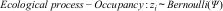
The observational process is modeled as a Bernoulli random variable conditional on the occupancy (z) and the probability of detection (p). The parameter p models the imperfect detectability of species due to trap type, abiotic conditions, or seasonal trends. In this way, the observational process is:
where, the observed variable yij is our measurement of occurrence at the site during the survey j (j=1,…,2, in our study). Detection probability, p, refers to all individuals inhabiting a site together.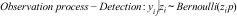
We constructed an a priori set of candidate models to evaluate whether fire or vegetation structure affect the occupation probability. As covariates we used the sites’ treatment or condition (burned or unburned), vegetation structure described by the first two components of the PCA, sampling occasion (1–4), potential food availability (the number of edible plant species), and woody debris (percentage of woody debris at each site). All the covariates, except vegetation structure (pca1 and pca2), were standardized with mean 0 and standard deviation 1. Since we did not find differences between forest edge and forest interior in both treatment transects, we combined this data for analysis of fire effects on occupation probability.
We followed an information-based approach to select the best candidate models. In this way, we used the corrected Akaike information criterion (AICc) to select the models that represented species detection and best occupancy while using the fewest parameters (Burnham and Anderson, 2002). We considered that models with ΔAICc>4 units had low support (Burnham and Anderson, 2002). Additionally, we estimated the goodness-of-fit (GOF) of the best-ranked models throughout parametric bootstrapping (Fiske and Chandler, 2011). Analyses were conducted using functions implemented in ‘ade4’ and ‘unmarked’ R packages (R Core Team, 2017).
ResultsFire effects on vegetation structureThe comparison of the Hill numbers revealed that fire has long-term effects on plant diversity at the Bojonawi Civil Society Reserve (Fig. 3A–C). Three years after the last fire event, the average richness of species at burned sites was lower than at unburned sites (Fig. 3A; t-test t=−6.93, p-value<0.0001). The same trend was observed when species abundances were considered to estimate the Hill numbers q=1 (Fig. 3B; t-test t=−5.81, p-value<0.0001) and q=2 (Fig. 3C; t-test t=−3.14, p-value<0.001). Fire also affected the distribution of vegetation strata (Fig. 3D). As indicated in the first component of the PCA (pca1), which explained 62.5% of the variance, fire mainly affected the ground, herbaceous, and arboreal strata. Unburned sites with a high coverage of ground and arboreal strata were on the positive side of the first component axis, while burned sites with high coverage of the herbaceous stratum were on the negative side. The second component (pca2), which explained 18.6% of the variance, described the variability of sites in terms of the shrub and low-arboreal strata. The distribution of sites along this component was not linked to fire effect.
 The first three Hill Numbers describing plant diversity at burned and unburned sites. Contour of violins plots provide information about the probability density of the data, boxplots within violins show median, quartiles and range. (D) Principal components analysis (PCA) that summarizes the information of the vegetation structure estimated at the study sites. Habitat variables herb=herbaceous, SubArb=subarboreal, Arb=arboreal. The biggest circle and triangle represent the centroid position in the ordination plane of burned and unburned sites.")
Vegetation structure and plant diversity at the Bojonawi Civil Society Reserve. (A–C) The first three Hill Numbers describing plant diversity at burned and unburned sites. Contour of violins plots provide information about the probability density of the data, boxplots within violins show median, quartiles and range. (D) Principal components analysis (PCA) that summarizes the information of the vegetation structure estimated at the study sites. Habitat variables herb=herbaceous, SubArb=subarboreal, Arb=arboreal. The biggest circle and triangle represent the centroid position in the ordination plane of burned and unburned sites.
A sampling effort of 10,560 trap/nights resulted in the capture of 134 individuals, comprising six species: two rodents and four marsupials. We captured 49 individuals of Zigodontomys brevicauda (cane mouse), 67 of Oecomys sp. (arboreal rice rats), 15 of Didelphis marsupialis (common opossum), 1 of Philander cf. opossum Linnaeus, 1758 (gray four-eyed opossum), 1 of Marmosops cf. noctivagus Tschudi, 1845 (white-bellied slender opossum), and 1 of Marmosa sp. Gray, 1821. We detected the first three species in the four sampling periods, while Marmosops cf. noctivagus, Philander cf. opossum, and Marmosa sp. were detected only once during the sampling of March 2020 in the unburned area. Oecomys sp. and D. maruspialis were detected in both burned and unburned areas, but their abundance peaked in the unburned sites (Fig. 4A and B). Zigodontomys brevicauda was primarily found in burned areas (Fig. 4C). We did not found differences between forest edge and forest interior for these three species in both treatments (Fig. 4). We recorded 31 recaptures of 19 different individuals, mainly of Z. brevicauda, captured during the sampling season of March 2020.

Average number of individuals per sampling recorded in burned and unburned areas in Bojonawi Civil Society Reserve. At each sampling, 240 Sherman traps were set in unburned and burned forest and opened for 11 nights in a row. Bars represent 1SD. Results displayed comprise 4 samplings periods.
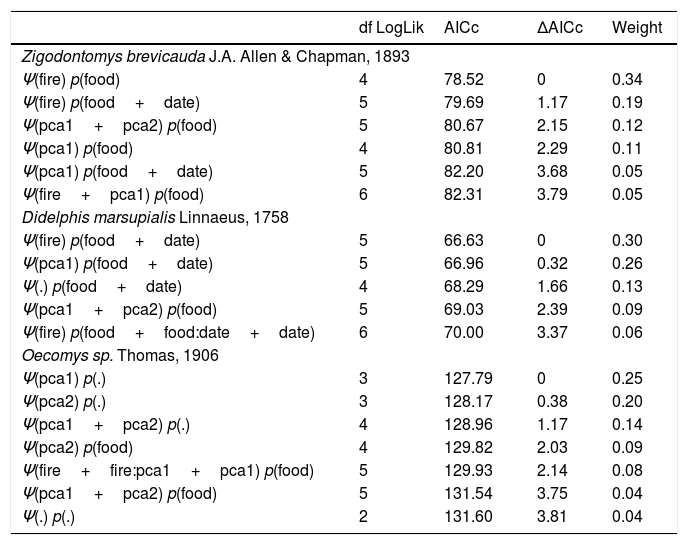
Parametric bootstrapping showed that all the best-ranked models fit the data well. The effect of fire was included among the most plausible models for Z. brevicauda and D. marsupialis (Table 1). Model predictions showed that the occupancy probability changed between burned and unburned sites. Z. brevicauda had the broadest distribution across burned sites (Fig. 5a), while D. marsupialis had the broadest distribution across unburned sites (Fig. 5D). The differences in vegetation structure linked to fire (‘pca1’) were also included in some of the most plausible models: both species had a higher probability of occupancy in sites with a greater frequency of the arboreal strata (Fig. 5B and D). For both species, the detection covariates included among the best-ranked models were the sampling occasion and potential food availability (i.e. number of species that potentially provide food) (Table 1). The probability of detection increased during the last sampling periods (Fig. 5E). Meanwhile, the probability of detection for Z. brevicauda and D. marsupialis decreased at sites with higher potential food availability provided by the vegetation (Fig. 5F).
Occupancy models ranked according to their AICc values. AICc indicate the empirical support for each model, relative to others. We only show the models with ΔAICc<4 and p-value>0.01 in a bootstrapped goodness-of-fit test on detection history frequencies. p=detection probability; Ψ=occupation probability; pca1=gradient of; pca2=gradient of.; fire=burned forest; food=the number of potential edible plant species; date=sampling occasion.
| df LogLik | AICc | ΔAICc | Weight | ||
|---|---|---|---|---|---|
| Zigodontomys brevicauda J.A. Allen & Chapman, 1893 | |||||
| Ψ(fire) p(food) | 4 | 78.52 | 0 | 0.34 | |
| Ψ(fire) p(food+date) | 5 | 79.69 | 1.17 | 0.19 | |
| Ψ(pca1+pca2) p(food) | 5 | 80.67 | 2.15 | 0.12 | |
| Ψ(pca1) p(food) | 4 | 80.81 | 2.29 | 0.11 | |
| Ψ(pca1) p(food+date) | 5 | 82.20 | 3.68 | 0.05 | |
| Ψ(fire+pca1) p(food) | 6 | 82.31 | 3.79 | 0.05 | |
| Didelphis marsupialis Linnaeus, 1758 | |||||
| Ψ(fire) p(food+date) | 5 | 66.63 | 0 | 0.30 | |
| Ψ(pca1) p(food+date) | 5 | 66.96 | 0.32 | 0.26 | |
| Ψ(.) p(food+date) | 4 | 68.29 | 1.66 | 0.13 | |
| Ψ(pca1+pca2) p(food) | 5 | 69.03 | 2.39 | 0.09 | |
| Ψ(fire) p(food+food:date+date) | 6 | 70.00 | 3.37 | 0.06 | |
| Oecomys sp. Thomas, 1906 | |||||
| Ψ(pca1) p(.) | 3 | 127.79 | 0 | 0.25 | |
| Ψ(pca2) p(.) | 3 | 128.17 | 0.38 | 0.20 | |
| Ψ(pca1+pca2) p(.) | 4 | 128.96 | 1.17 | 0.14 | |
| Ψ(pca2) p(food) | 4 | 129.82 | 2.03 | 0.09 | |
| Ψ(fire+fire:pca1+pca1) p(food) | 5 | 129.93 | 2.14 | 0.08 | |
| Ψ(pca1+pca2) p(food) | 5 | 131.54 | 3.75 | 0.04 | |
| Ψ(.) p(.) | 2 | 131.60 | 3.81 | 0.04 | |
 and D. marsupialis (D–F) according to the two best-ranked models in Table 1. Values of the multivariate gradient of vegetation structure were extracted from Axis 1 of the principal component analysis. The lowest negative values indicated the highest coverage of the herbaceous stratum, whereas the highest positive values indicated the highest coverage of ground and arboreal strata.")
Occupancy and detection probability of Z. brevicauda (A–C) and D. marsupialis (D–F) according to the two best-ranked models in Table 1. Values of the multivariate gradient of vegetation structure were extracted from Axis 1 of the principal component analysis. The lowest negative values indicated the highest coverage of the herbaceous stratum, whereas the highest positive values indicated the highest coverage of ground and arboreal strata.
The occupancy probability of Oecomys sp. did not differ between burned and unburned sites but was mainly explained by the vegetation structure. Both principal components, pca1 and pca2, were included as covariates of the best-ranked models (Table 1). The predicted occupancy probability increased in the positive sides of the pca1 and pca2 (Fig. 6A and B), suggesting that Oecomys sp. distribution increased at sites with greater coverage of shrub and arboreal strata (Fig. 6A and B), accordingly with the biology and ecology of this species (Emmons and Feer, 1997; Hershkovitz, 1960). The best-ranked models did not include a covariable explaining the detection probability of this species.

Occupancy and detection probability of Oecomys sp. according to the two best-ranked models in Table 1. Values of the multivariate gradient of vegetation structure were extracted from the first two axes of the principal component analysis. In both axes, the highest positive values indicate the highest coverage of ground and arboreal strata.
Our findings showed that fires in gallery forests have a strong impact on the community structure of non-volant small mammals, even three years after the last fire event. The most interesting result was that the effect of fire was not consistent among species, so that we found that fire may restrict (D. marsupialis), promote (Z. brevicauda) or have no impact (Oecomys sp) on the occurrence of mammal species. Such findings might reflect the differential effect of fire on vegetation strata. Previous studies have shown that vegetation's vertical stratification constrains tropical mammal species’ occurrence since each stratum provides distinct resources and retreats, such as food or protection against predators (Camargo et al., 2018; Chia et al., 2016; Pereira et al., 2010; Vieira and Monteiro-Filho, 2003). In line with these studies, our models suggest that fire has a more substantial effect on those species using the strata that disappeared because of fire.
According to our models, D. marsupialis was the species more affected by fire in the mid-term. The model predicted that its occurrence was affected by fire, potential food availability, and the sampling period (Table 1 and Fig. 5). Previous studies found that moderate-to-severe fires reduce the availability of food resources and structural habitat components at the understory (Bassett et al., 2015; Chia et al., 2016). Considering that D. marsupialis feeds mainly on plants, invertebrates and small vertebrates associated with the understory (Cáceres, 2004; Cerqueira and Tribe, 2008; Julien-Laferriere and Atramentowicz, 1990), our model suggest that the marked low abundance of the species in burned forests might respond to the structural downgrading of the understory because of fire. Such a response may be generalizable to other Didelphis species, because they exploit the structural components of the understory in a similar way. Didelphis albiventris Lund (1840), for instance, mainly inhabits the understory and canopy strata in gallery forests (Camargo et al., 2018).
Contrary to D. marsupialis, the occurrence of the cane mouse (Z. brevicauda) was positively affected by the last fire event. The interactive effect of fire, vegetation structure, and potential food availability suggests that fire increases the occurrence of Z. brevicauda by increasing the extent of open habitats, such as savannas, shrubs, grasslands, or crops (Voss, 1991). In comparison with the study's unburned sites, the burned sites were in an early successional stage with less complex vegetation composition and structure, less plant diversity and abundance, and greater dominance of ground and herbaceous strata. Such modifications can lead to a depletion of resources and to a reduction of habitats for specialist species, but may favor those species with omnivorous diets, flexible life history traits, and generalists habits (Kelly et al., 2010). In fact, Z. brevicauda has a generalist diet that includes insects and plants (Voss, 1991), and its dominance in burned forests may be partially explained by the higher insect availability during the first successional stages in areas affected by fires (Briani et al., 2004; Vasconcelos et al., 2009; Vieira and Briani, 2013). Likewise, this pattern has been reported for other species in the Cricetidae family such as Calomys tener Winge, 1887 and Calomys expulsus Lund, 1841 (Vieira and Briani, 2013). These patterns were also found in forests of the Brazilian Cerrado (Camargo et al., 2018) and the Amazonian forests (Barlow and Peres, 2006), where fire favored the invasion of generalist species adapted to open and altered areas.
The common mouse (Oecomys sp.) was found in both burned and unburned forests, and our models suggest that fire does not affect its occurrence although it was more frequent in the unburned areas. Rather, its occupancy was influenced by the structure of the vegetation and thus, the relationship between Oecomys sp. occupancy and fire ocurrence was not entirely conclusive. Oecomys distribution increased at sites with large shrub and extensive arboreal strata (Table 1 and Fig. 6), probably due to the specific habitat requirements of this species. A previous study showed that Oecomys nests in taller trees, epiphytes or vines (Hershkovitz, 1960) and that its diet strongly depends on fruits and seeds of shrubs and trees during the wet and dry periods (Emmons and Feer, 1997; Hershkovitz, 1960). Moreover, previous trophic ecology assessments of non-volant small mammals in El Cerrado, Brazil, found that Oecomys cleberi, an arboreal forest specialist that feed on fruits and seeds (Paglia et al., 2012), had a trophic specialization with a diet based on fruits (Ribeiro et al., 2019). The lack of a differential response to fire might be due to the availability of resources provided by higher strata vegetation in both burned and unburned areas, demonstrating that this species can thrive in postfire surroundings.
Although Oecomys sp. was the community's dominating species from the unburned forest, we also registered marsupials such as Marmosops cf. Noctivagus, Marmosa sp. And Philander cf. opossum. The occurrence of these marsupials, absent in the burned areas, is usually associated with primary and secondary humid forest ecosystems (Cuartas-Calle and Muñoz-Arango, 2003; Emmons and Feer, 1997), where they inhabit trees and fallen logs in the lower strata of the forest, feeding on fruits, invertebrates and small vertebrates (Cuartas-Calle and Muñoz-Arango, 2003; Emmons and Feer, 1997). As fire primarily affected arboreal strata, the impact seemed to be more intense on these marsupials, leading to their local extinction in the burned areas. However, we have to take with caution these results as such species were captured in low numbers.
Overall, our findings demonstrated that fires may structure the non-volant small mammal communities, with possible negative effects on the functioning of gallery forests. Our results show the strong impacts of a fire event in different small-mammal populations, but further studies involving fire event replicas are needed to profound on the role of fire in structuring of tropical communities. The forests of South America have higher probabilities of facing adverse climatic conditions with less resistance to climatic stress (Anjos and de Toledo, 2018) and less capability of adapting and recovering after disturbances, especially when these forests are compared with other types of vegetation such as natural savannas (Anjos and de Toledo, 2018). The low recovery capability of the forests exerts a direct influence on the faunal communities, that may result in cascading effects, disrupting the cycle of natural regeneration of plants and the ecosystem functioning (Bellard et al., 2012; Campos et al., 2017; Malhi et al., 2014).
ConclusionsTo our knowledge, this is the first study exploring the effects of fire on non-volant small mammals in neotropical savanna-gallery forests of northern South America, and it enhances our knowledge about the effects of fire on neotropical fauna. Overall, our results showed that vegetation attributes have a fundamental role in the occupancy by small mammals. Species associated with close canopies were negatively influenced by fire, but species associated with open habitats had high representation in burned areas. Such findings suggest that fire can modulate habitat characteristics through its influence over vegetation structure and composition.
Our study also reinforces the importance of wildfire prevention in fire-sensitive ecosystems like the Colombo-Venezuelan llanos’ gallery forests. Tropical forests are not adapted to resist disturbances such as fire; thus, a gallery forest's ecology can be affected even by a low-intensity fire, and the effects extend for several years after the fire event. Our findings suggest that fire might disrupt gallery forests’ functioning by affecting keystone plant-animal interactions that drive ecosystem functions such as seed predation and dispersal. It is critical to continue monitoring for understanding fire effect in the long term. Post-fire faunal successional trajectories are still unknown in the tropics.
FundingThis research was funded by Colciencias 757 National Doctorate Scholarship, by Universidad Nacional de Colombia through the Convocatoria Nacional para el Apoyo a Proyectos de Investigación y Creación Artística 2017–2018, by the Grupo de Investigación en Ecología del Paisaje y Modelación de Ecosistemas – Ecolmod, through the support by NAS Subaward Letter No. 2000007526, PEER Cycle 5 funding, the American Society of Mammalogists through the Latin American student field grant 2017, The Rufford Foundation – Small Grant, and the L’Oréal-UNESCO For Women in Science Colombia 2018 scholarship. We wish to thank Idea wild and Neotropical Grassland Conservancy for the donated equipment.
Authors’ contributionTMG and DA conceived and designed the study; TMG, DA and AM designed the methodology; TMG and DA collected the data; TMG and JDGT analyzed the data; TMG led the writing with inputs from DA, AM and JDGT. All authors contributed critically to the manuscript drafts and gave their final approval for publication.
We thank Professor Pedro Sanchez and La Palmita Foundation for lending us the Sherman traps. The field and laboratory work for this project was intense and the help and support of the following people was essential: Francisco Luque, María Constanza Meza, Eduardo Molina, Nathalia Moreno, Catherine Mora, Javier Alejandro Salas, Francisco Castro, Alejandra Reyes Palacios, Gina Sierra, Brayan Marín, Beyker Castañeda, Henry Esteves, Nelcy Vega, Jacinto Terán, Catalina Cárdenas, Darwin Morales, and Miguel Rodríguez. We would like to thank Fundación Omacha and particularly its scientific director Fernando Trujillo for their continuous support and permission to work in the Bojonawi Civil Society Reserve. We also thank Thomas Richard Defler for the revision of the manuscript. The research was done under the scientific research permit no. Resolution 0255 Date March 14, 2014 of the Autoridad Nacional de Licencias Ambientales (ANLA). This research was approved by the Ethical Committee of the Sciences Faculty of Universidad Nacional de Colombia, Bogotá, Nos. 02-2018 and 07-2019. Finally, we wish to thank to the reviewers for their comments and suggestion that greatly improved the manuscript.

