



Burmese pythons are established in the Everglades and are expanding their population in Florida to more urbanized and fragmented habitats. Understanding movement and dispersal behavior contributes to our knowledge of how landscape factors influence the persistence of pythons in Florida's habitat. Our goal was to examine personality-dependent dispersal in juvenile Burmese pythons by creating behavioral scenarios of risk-taking behavior on a shy–bold continuum using an individual-based model. We observed that a behaviorally plastic strategy best resembled empirically derived patterns of the Burmese python's expansion from the Everglades into the increasingly urbanized landscapes of Homestead and Miami, Florida. This result is consistent with the notion that animal personalities can be flexible in different situations and that animals must make decisions based on trade-offs while dispersing.
Invasion biology has a long tradition of identifying traits that could explain between-species dissimilarities in species’ abilities to succeed as invaders (Cote et al., 2010). However, examining the average behavioral response of a population as a whole masks the variation between individuals that likely drives invasion dynamics, particularly those characteristics that may only be advantageous in certain phases of the invasion. The process of an invasion is composed of several stages starting with initial introduction and spread and ending with integration into the ecological community (Vermeij, 1996). Researchers are increasingly focusing on different behaviors that help invaders transition from one stage to another while recognizing that these behaviors may not be as beneficial to the persistence of the invasive population in the next phase of invasion (Cote et al., 2010).
Personality-dependent dispersal, where personality types such as boldness, aggressiveness, and sociability are linked to the propensity to disperse, is particularly relevant in studying the spread of invasive populations (Duckworth and Badyaev, 2007; Sih et al., 2012). The net movement of the most dispersive individuals of a population determines its rate of expansion, even when long-distance dispersal events are rare (Neubert and Caswell, 2000; Bartón et al., 2012). Fraser et al. (2001) demonstrated that movement behavior is heterogeneous within a population, thus intraspecific differences in demography, behavior, or personality are important for describing dispersal kernels for an invading population. For example, western bluebirds (Sialia mexicana) on an expanding front across the western United States were more likely to be aggressive and to thus outcompete sister taxa; however, individuals behind the front in the established range were more likely to be less aggressive because aggressiveness was correlated with poor parental care in males (Duckworth and Badyaev, 2007). Invasive cane toads (Chaunus [Bufo] marinus) in Australia employed a range of sociality depending on their position along the colonization front (González-Bernal et al., 2014). Boldness has been linked to dispersal tendency in a variety of species (e.g., Lepomis gibbosus, Coleman and Wilson, 1998; Parus major, Dingemanse et al., 2003; Vulpes velox, Bremner-Harrison et al., 2004; Neogobius melanostomus, Myles-Gonzalez et al., 2015).
Boldness is the tendency of organisms to explore and move through unfamiliar space and novel situations (Wilson et al., 1993). Bold individuals tend to move greater distances and to be riskier in how they explore unfamiliar landscapes and in their antipredator response (Bartón et al., 2012). While bolder dispersers move greater distances, they also have higher probabilities of mortality (Azevedo and Young, 2006). For example, the boldest reintroduced swift foxes in Montana moved the farthest from their release sites but experienced lower survival compared to individuals who limited their movements (Bremner-Harrison et al., 2004). Clearly, behaviors that maximize an individual's dispersal distance may not be advantageous to individual survival (Bartón et al., 2012). This suggests that the expansion of a population is driven by within-individual variation in balancing risk and dispersal distance (Zollner and Lima, 2005), or that boldness varies between individuals with those on the expanding front bolder than the average individual in the core population and the rare, bold survivor driving the expansion (Fraser et al., 2001; Bartón et al., 2012).
Among the most significant biological invasions currently taking place in the eastern United States is the Burmese python in southern Florida. Pythons are well established in the Everglades, but they appear to be expanding their range northward into more urbanized and heterogeneous landscapes (Dorcas and Willson, 2011). While south Florida's habitat is ideal for the python (e.g., abundant prey, similar climate as native range), it is not without risks of mortality (e.g., predators, road networks, machinery).
Understanding resource distribution and habitat suitability as well as movement and behavior is important in predicting the python population's ability to persist in novel, fragmented habitats outside of the Everglades (Taylor et al., 1993; Knowlton and Graham, 2010). Dispersal and movement are recognized as key processes involved in measuring landscape connectivity for particular species (Taylor et al., 1993). However, little is known about the behavior and movement of dispersing juvenile pythons due to their evasiveness and their use of remote habitat. Because of the lack of information on Burmese python dispersal behavior, there is little direct evidence that Burmese pythons display personality types; however, the pervasiveness of behavioral syndromes in well-studied species suggests a high likelihood that Burmese pythons display consistent differences in behavior related to movement and exploration (Sih et al., 2004). Preliminary research on boldness in Burmese pythons indicates an effect of relatedness on risk-taking strategy in laboratory trials (Pittman, unpubl. data). Additionally, Pittman et al. (2014) found that pythons have sophisticated navigational abilities which could contribute to behavioral decisions during dispersal. Studying the behavioral components of how pythons move through Florida's landscape thus contributes to our understanding of their ability to spread into novel habitat. Additionally, knowledge of patterns of dispersal aids our ability to plan control methods that could manage the spread of Burmese pythons and other invasive species in the state.
Our objective was to use a spatially explicit individual-based model (IBM) to investigate boldness on the expanding front of the Burmese python population in southern Florida as the population expanded from the Everglades into human-dominated landscapes. IBMs can illustrate the interaction between individual behaviors and landscape configuration and characteristics, an interaction that drives animal movement behavior (Zollner and Lima, 1999). Our goal was to simulate individual behaviors on the edge of the population so we could observe the rare dispersal events that drive a population's expansion. We predicted that individuals on the front characterized as bold would move faster and farther than shy individuals, and that the rate of expansion of bold individuals would most closely resemble the rate observed in the field. We also predicted that the final range occupied by pythons as facilitated by bold dispersers would most closely resemble the observed range during this same time frame. We expected bolder individuals to experience higher rates of mortality and establish home ranges farther away from their release locations in comparison to shyer individuals.
Material and methodsModel overviewOur goal was to simulate risk-taking behavior in dispersing juvenile Burmese pythons on the leading edge of an expanding front. We created 6 behavioral scenarios on a shy–bold scale and modeled dispersal of 25 virtual pythons per scenario per dispersal season. Each scenario ran for a dispersal season of 6 months over a 10-year period from 2004 to 2013. The Early Detection & Distribution Mapping System (EDDMapS) database provided extensive python presence data recorded over 10 years (2004–2013) that we used to pattern-match (Grimm and Railsback, 2013) our model outputs (see Appendix A, Model study system and Main Text, Analysis). We only modeled individuals on the leading edge of the front; at the beginning of each dispersal season, we determined the new leading edge and selected a new random sample of 25 individuals. Thus, we were able to simulate python expansion across our study site while restricting our focus to virtual snakes on the leading edge of the population.
Modeling frameworkWe used the spatially explicit individual-based model Spatially Explicit Animal Response to Composition of Habitat (SEARCH). SEARCH simulates animal dispersal and home-range establishment on a virtual landscape with a high degree of behavioral complexity (Pauli et al., 2013). The program interfaces with ArcGIS (ESRI, Redlands, CA, USA) to build a virtual landscape, which is comprised of vector-based maps representing animal movement, foraging opportunities, risk of mortality, habitat suitability, and exclusive occupancy of resident animals (Pauli et al., 2013; Table 1). Each map contains specific field definitions based on different GIS classifications, and virtual animals individually alter their behavior or physiological state based on these parameters (Pauli et al., 2013). Dispersers can be introduced to the landscape via a point release map or may be “born” on the landscape through the reproduction of resident animals (Pauli et al., 2013). Each individual generates its own memory map. This represents the information it perceives from its environment and is used in making settlement decisions. Virtual animals respond to per timestep mortality and energetics and change behavioral states (e.g., searching vs. foraging, risky vs. safe) as they interact with the landscape (Pauli et al., 2013; Table 1). Parameters governing behavior, energetics, home-range requirements, and resident reproduction can be modified to include heterogeneity in animal response caused by sex, time, and behavioral state (Pauli et al., 2013; Blythe et al., 2011). Appendices are available in the supplementary online material. See Appendix A for a full description of the model study system, map inputs, and model parameterization and the word box for details on behavioral scenarios.
Simulation parameters coded into the vector-based risk map's attribute table and read by the individual-based model SEARCH. Risk parameters were parameterized based on 2014–2015 data from a radio telemetry field study.
| Land cover type | Description | Probability of mortality per timestep (12-h) |
|---|---|---|
| Human-dominated habitat | Correspond to the following land cover types defined by the movement map: agricultural lands, urban landscapes, and canals. These areas are heavily influenced by human activities. | 0.0019 |
| Naturally-dominated habitat | Correspond to the natural habitat cover type defined by the movement map. These areas have less anthropogenic influence in comparison to altered habitats. | 0.0053 |
We simulated dispersal and home range establishment of juvenile pythons on an agricultural and urban interface in southern Florida between the southeastern Everglades, Homestead, and south Miami (Fig. 1 and Table 1). Southern Florida is located in a subtropical climate characterized by a wet and a dry season. This section of the Everglades is comprised of freshwater sloughs, marl prairies, tropical hardwood hammocks, and pinelands. Agricultural lands and low-density urban development characterize Homestead, Florida, while urbanization intensity increases rapidly as Homestead connects to southern Miami and approaches the city center. These areas are anthropogenically connected via a dense road network and canal waterways. We chose this study area because it was comprised of land cover types from which we had empirical data on juvenile python movement (Table 1; Appendix A, Model study system). See Appendix B for a full description of why and how we chose this modeling extent.
Analysis
We used a pattern-oriented approach to compare model outputs to an empirical presence-only dataset. Patterns are viewed as foundational to the structure and processes of a system; therefore, if a particular pattern observed empirically is not observed in a particular modeling scenario, that model is presumed to perform unrealistically and can be discarded (Semeniuk et al., 2012).
We compared the simulated rate of expansion from each scenario to the observed spread of the python population. We estimated an index of the rate of expansion of the population's distribution in the empirical data and the output of each simulation scenario (Appendix C). To pattern-match, we compared the index of the rate of expansion from each simulation scenario to the empirical dataset to determine which behavioral type best represented dispersing pythons in southern Florida (Appendix C). To match a second pattern we compared the range occupied by Burmese pythons in the empirical dataset to the range occupied by virtual pythons in each behavioral scenario (Appendix C). We defined 4 categories to compare: Non-invaded area (includes areas not invaded through 2013), Phase 1 (the combined ranges from 2004 to 2007), Phase 2 (the combined ranges from 2008 to 2010), and Phase 3 (the combined ranges from 2011 to 2013). We chose these categories over comparing the range for each incremental year in order to focus the map comparison on the location of the leading edge instead of pixel-to-pixel agreement of each year's range.
ResultsSummary statisticsWhen interpreting the results across the gradient of behaviors, we generally use the term “bolder” to refer to individuals on the bold end of the shy–bold continuum, and “shyer” to refer to those on the shy end of the continuum.
Bolder virtual pythons traveled greater distances and moved farther from their release locations than shyer virtual pythons (Fig. 2A and B). Shyer and bolder virtual pythons switched infrequently between modes (Fig. 2C) with shyer pythons spending most of their time in safe mode and bolder pythons in risky mode (Fig. 2D). Behaviorally plastic animals switched frequently between safe and risky modes, and spent about half of their time in each mode (Fig. 2C and D). However, overall intermediate animals did not switch frequently between modes; they remained in one mode for most of their dispersal period with a bias toward remaining in risky mode (Fig. 2C and D).
 mean total distance traveled, (B) mean straight-line distance traveled, (C) mean number of switches between modes, (D) mean proportion of time spent in risky mode, (E) mean number of timesteps, and (F) proportion of fates, across all individuals within each behavioral scenario.")
Summary of the (A) mean total distance traveled, (B) mean straight-line distance traveled, (C) mean number of switches between modes, (D) mean proportion of time spent in risky mode, (E) mean number of timesteps, and (F) proportion of fates, across all individuals within each behavioral scenario.
Shyer virtual pythons moved for a greater number of timesteps in comparison to bolder pythons (Fig. 2E). A greater proportion of shyer pythons were able to establish home ranges while a greater proportion of bolder pythons died due to riskier behavior (Fig. 2F). Behaviorally plastic and intermediate animals experienced high mortality as well, with greater than 50% subjected to mortality (Fig. 2F).
Shyer virtual pythons that traveled the farthest distance from the prior year's leading edge were more likely to be alive at the end of the dispersal season in comparison to bold pythons, who were more likely to have died (Fig. 2). The most dispersive behaviorally plastic pythons also were more likely to have had died by the end of the season. Accordingly, the most dispersive shy animals from one year were more likely to determine the next year's leading edge while the most dispersive bold animals were more likely to have died and have no contribution to the next year's leading edge.
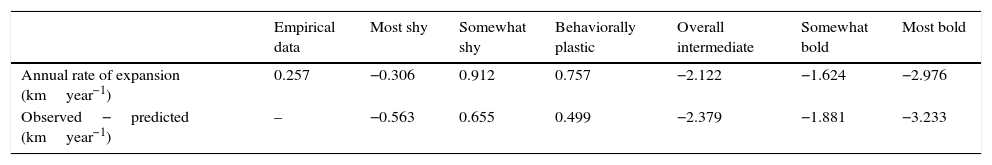
Pattern matchingThe empirically derived index of the annual rate of expansion of the Burmese python population showed a rate of 0.257kmyear−1 (Table 2). While the somewhat shy behavioral scenario also showed a positive rate of spread, the behaviorally plastic scenario most closely resembled the empirically derived index, only differing by 0.499kmyear−1. In contrast, the bolder scenarios yielded negative rates of spread: the rate decreased by nearly 6.5× for the somewhat bold scenario and by 11.5× for the most bold scenario.
Index of the annual rate of expansion of each behavioral scenario compared to the empirically-derived index of rate of expansion.
| Empirical data | Most shy | Somewhat shy | Behaviorally plastic | Overall intermediate | Somewhat bold | Most bold | |
|---|---|---|---|---|---|---|---|
| Annual rate of expansion (kmyear−1) | 0.257 | −0.306 | 0.912 | 0.757 | −2.122 | −1.624 | −2.976 |
| Observed−predicted (kmyear−1) | – | −0.563 | 0.655 | 0.499 | −2.379 | −1.881 | −3.233 |
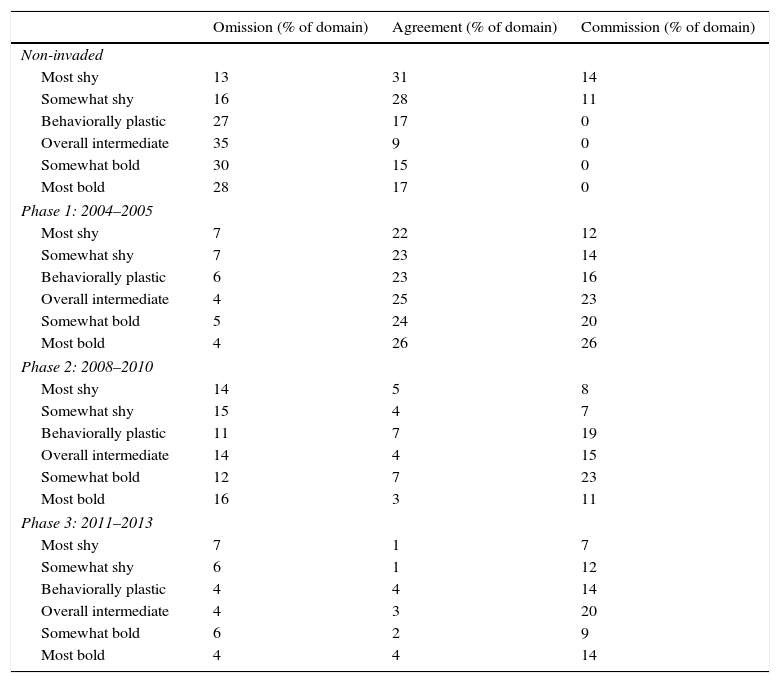
Overall, the simulated ranges had low overall map agreement with the empirical range (Table 3). Agreement was highest between non-invaded and Phase 1 pixels, but it decreased over time from Phase 1 through Phase 3 (i.e., from 2004 to 2013). Agreement was consistently high for each phase between the empirical range and the behaviorally plastic scenario's range relative to the other scenarios. Agreement within each phase was also second highest for bolder behavioral scenarios. However, the shyer behavioral scenarios had greater overall agreement because they had the greatest agreement with non-invaded area.
Map agreement and omission and commission errors between the empirically derived geographical extent of the Burmese python population's range and the 6 simulated behavioral scenarios’ ranges.
| Omission (% of domain) | Agreement (% of domain) | Commission (% of domain) | |
|---|---|---|---|
| Non-invaded | |||
| Most shy | 13 | 31 | 14 |
| Somewhat shy | 16 | 28 | 11 |
| Behaviorally plastic | 27 | 17 | 0 |
| Overall intermediate | 35 | 9 | 0 |
| Somewhat bold | 30 | 15 | 0 |
| Most bold | 28 | 17 | 0 |
| Phase 1: 2004–2005 | |||
| Most shy | 7 | 22 | 12 |
| Somewhat shy | 7 | 23 | 14 |
| Behaviorally plastic | 6 | 23 | 16 |
| Overall intermediate | 4 | 25 | 23 |
| Somewhat bold | 5 | 24 | 20 |
| Most bold | 4 | 26 | 26 |
| Phase 2: 2008–2010 | |||
| Most shy | 14 | 5 | 8 |
| Somewhat shy | 15 | 4 | 7 |
| Behaviorally plastic | 11 | 7 | 19 |
| Overall intermediate | 14 | 4 | 15 |
| Somewhat bold | 12 | 7 | 23 |
| Most bold | 16 | 3 | 11 |
| Phase 3: 2011–2013 | |||
| Most shy | 7 | 1 | 7 |
| Somewhat shy | 6 | 1 | 12 |
| Behaviorally plastic | 4 | 4 | 14 |
| Overall intermediate | 4 | 3 | 20 |
| Somewhat bold | 6 | 2 | 9 |
| Most bold | 4 | 4 | 14 |
Our IBM supported our predictions of how personality-dependent dispersal can impact the spread of an expanding population. Along the shy–bold continuum, bolder virtual snakes spent a greater proportion of time in risky mode. While spending more time in risky mode resulted in greater dispersal distances, it also resulted in higher rates of mortality. This meant that the individuals who traveled the farthest in the bolder scenarios did not always contribute to the annual advancement of the front since they would not survive to the end of the dispersal season. Even so, the bolder survivors dispersed consistently with our expectations since the expanding edge still advanced farther across the modeling extent in the bolder behavioral scenarios. Additionally, while overall map agreement was low between the simulated range and the empirical range, this outcome was expected because our simulated population size was much smaller than the actual population size (and therefore we did not expect high map agreement). We were therefore most interested in consistency between the behavioral scenario most closely matching our index of expansion and the behavioral scenario with the highest map agreement.
We expected the most bold scenario to be the best match with our empirically observed patterns of annual rate of spread and the geographical extent of the python population. Instead, the behaviorally plastic scenario was the best fit based upon its closest match to the observed rate of expansion and the population's geographical extent. Although the two shyer behavioral scenarios had higher agreement with the empirically observed geographical extent, a large proportion of the percent agreement is due to matching the non-invaded area (Table 3). For Phases 1 through 3, (i.e., from the years 2004–2013), the shyer scenarios had the lowest agreement in comparison to the other behavioral scenarios. The behaviorally plastic scenario has the third highest overall agreement with the empirical dataset and consistently had the first or second highest agreement with Phases 1 through 3. Coupled with the best match for the annual rate of the python population's expansion, our results suggest that pythons with adaptively flexible dispersal behavior are more likely to be driving the expansion of the population into south Florida's urbanized landscapes than individuals that consistently display bold or shy behavioral phenotypes. This is further supported by the relative performance of the shyer and bolder scenarios simulated during different phases of the expansion: shyer snakes produced population extents similar to empirical observations in earlier phases while bolder snakes generated extents more similar overall to empirical observations relative to the shyer snakes. This demonstrates the success of dichotomous behavior within the same population at different stages of expansion.
Although animal personality suggests consistency of behaviors, animal personality and behavioral plasticity do not exist separately from one another (Sih et al., 2004). There is increasing evidence that animals can be relatively consistent in their behaviors while still retaining flexibility (Briffa et al., 2008). Animals may adjust their personality-dependent behaviors depending on social situations (Natarajan et al., 2009), predation risk (Quinn and Cresswell, 2005), and environmental variables (Bonte et al., 2007). Fragmentation and human disturbance also impact landscape connectivity on an individual level: movement and dispersal can vary between individuals, among or within landscapes (Knowlton and Graham, 2010). Therefore, future IBMs on the Burmese python system would benefit from relating behaviors to specific landscape characteristics and environmental variables. This would lead to specific knowledge of how landscape factors drive behavioral plasticity in dispersing juveniles and contribute to our understanding of how Florida's heterogeneous landscape is able to support a viable population of Burmese pythons (Knowlton and Graham, 2010).
While bold individuals are more likely to explore novel situations, they are not necessarily better equipped to survive; rather, individuals with behavioral flexibility have better responses to novel conditions (Sih et al., 2004). The spread of a species may be best facilitated by populations comprised of variation in traits between dispersers on the leading edge, particularly when different behaviors specialize on different population densities and disperse according to these specializations (Fogarty et al., 2010). Thus, it is better for an individual's fitness to balance risk and dispersal plastically than to adhere to a fully consistent behavior. This is especially true when animals are dispersing through fragmented landscapes where risk is heterogeneous. For example, moving slowly and exhibiting anti-predator behaviors is beneficial when moving through risky matrix, but it is not as beneficial to an individual when moving through less risky habitat (Zollner and Lima, 2005). Jepsen and Topping (2004) observed that populations of roe deer (Capreolus capreolus) with behavioral plasticity had higher population sizes, population persistence, and abilities to cope with patchy landscapes. Burmese pythons in Florida similarly make trade-offs as they move through Florida's landscape: pythons may use canals to make long distance movements between suitable habitat patches, but canals are risky due to the prevalence of American alligators (Alligator mississippiensis).
Although our IBM did not support our initial prediction, our results do not contradict the literature supporting risky movement and boldness. We were not modeling an invasive population spreading from its point of introduction into suitable, homogenous landscapes. The population we modeled is best described as a core population pushing out of a naturally dominated landscape into an increasingly urbanized landscape. As established in the literature, behavioral plasticity is most beneficial to the persistence of a population in this situation (Jepsen and Topping, 2004). Further research in the Burmese python system is needed to elucidate our understanding of within-individual behavioral plasticity and the mechanism that may be driving the adaptive advantage for plastic behavioral syndromes. Future research focusing on Burmese python boldness behavior will provide insight into the extent to which pythons exhibit consistent differences in boldness behavior, and this simulation study highlights the need for a detailed understanding of movement behavior in forecasting invasion dynamics. Nonetheless, our result that behaviorally plastic individuals best matched empirical patterns implies a need for more flexible definitions of connectivity that emphasize the importance of behavior and recognize that its influence can be dynamic and circumstance-specific. These trends may be particularly true for invasive species spreading from naturally-dominated to human-dominated landscapes, such as in our Burmese python case study.
Word boxBehavioral scenariosIn order to examine risk-taking behavior of dispersing juvenile pythons, we created six behavioral scenarios representing a gradient of responses to risk on a shy–bold scale: most shy, somewhat shy, behaviorally plastic, overall intermediate, somewhat bold, and most bold (Table 2S). These behaviors can be implemented in SEARCH by manipulating criteria for dispersers’ tendency to switch from or remain in “risky” or “safe” modes and modifying movement behaviors and risk of mortality within these modes. Animals switch modes based on user-defined probabilities. We initially selected 3 probabilities (0.1, 0.01, and 0.001) to implement a 3×3 fully-crossed design, but model testing revealed that 3 of the combinations resulted in duplicate behaviors. Therefore, we focused our investigation upon the 6 combinations that best reflected the spectrum of shy to bold we wished to investigate. See Appendix A for a full description of each behavioral scenario.
Conflicts of interestThe authors declare no conflicts of interest.
We thank P. Andreadis, I. Bartoszek, and C. Ervin for providing data and insight into the development of the model. We thank C. Day and L. D’Acunto for assistance with model development and software troubleshooting. We thank J. Doucette for assistance with software and K. Kellner for assistance with data summarization. This work was partially funded by a Purdue University Knox Fellowship grant to H. Mutascio and the National Science Foundation Postdoctoral Fellowship in Biology Program Grant No. 1309144.
The following are the supplementary data to this article:

