




Cross-habitat movements are crucial for persistence of beneficial insects in agricultural landscapes; however, much remains unknown on how landscape structure affects the spillover of beneficial insects between crop and non-crop habitats. To estimate the effects of landscape structure on the spillover of beneficial insects we sampled predatory wasps in pairs of forest remnants and adjacent coffee plantations along a gradient of landscape composition and configuration. We used dissimilarity indices to estimate wasp spillover and we assumed that high dissimilarity means less flow (and thus less spillover) between forest and coffee habitats. We collected a total of 9847 wasps classified into 75 species and 23 genera. Wasp dissimilarity between habitats decreased with increasing forest cover in the surrounding landscape and did not respond to landscape diversity, edge density or pesticide usage. Our findings suggest that wasps forage in coffee plantations but seem to rely on forest remnants to find unmanaged nesting sites and a constant supply of resources that are not available in the agricultural matrix, and are neither in landscapes with high compositional diversity or edge density. Therefore, forest conservation and restoration should be incorporated in agro-environmental schemes designed to improve the spillover of beneficial insects and provision of ecosystem services within coffee farmlands.
Natural enemy and pollinator insects provide important ecosystem services for agriculture; however, crops are intensively managed and the occurrence of these insects within the agricultural matrix depends on the spillover from adjacent habitats (Haan et al., 2020). These beneficial insects often rely on non-crop habitats such as forests, meadows and grasslands in the surrounding landscape to find a permanent and diverse supply of food items, refuge from crop management, nesting sites and overwintering habitats (Tscharntke et al., 2012; Bartual et al., 2019). Beneficial insects can also rely on landscape diversity, foraging in multiple crop and non-crop habitat types, to fulfill resource requirements and complete their life cycle (Tscharntke et al., 2012; Sirami et al., 2019). Landscape metrics related to landscape configuration such as habitat connectivity and edge density also influence the spillover of beneficial insects and how far they can disperse into an agricultural matrix (Haan et al., 2020).
Cross-habitat spillover between crop and non-crop habitats is not unidirectional and the flux of organisms can occur from natural habitats to crops and vice versa (Sackett et al., 2009; Tscharntke et al., 2012). Multiple factors can determine the magnitude and direction of spillover in agricultural landscapes including crop management, and high contrast in habitat structure and primary productivity between crops and natural habitats (Frost et al., 2015). Natural habitats are unmanaged, provide a constant supply of resources for biodiversity and often act as a source of beneficial insects and ecosystem services to adjacent agroecosystems (Bianchi et al., 2006; Tscharntke et al., 2012). In contrast, the pulses of resource availability in crops (e.g., pest outbreaks and mass flowering) can make the agricultural matrix a temporary source of beneficial insects to adjacent natural habitats (Rand et al., 2006; Frost et al., 2015; Madeira et al., 2016). Such cross-habitat movements are crucial for persistence of beneficial insects in agricultural landscapes where many species rely on different resources that are spatially separated in different crop and non-crop habitats (Tscharntke et al., 2012).
Recent studies conducted in Brazilian coffee-producing landscapes provide evidence that Atlantic Forest is a crucial habitat for insect conservation, and that high forest cover and proximity enhance the occurrence of beneficial insects into adjacent crops, resulting in high coffee fruit set and low levels of pest infestations (Saturni et al., 2016; Hipólito et al., 2018; Aristizábal and Metzger, 2019; Medeiros et al., 2019a, 2019b, González-Chaves et al., 2020). These studies estimated how landscape structure affects the occurrence of beneficial insects and provision of ecosystem services within coffee plantations; however, further investigation is required to better understand how landscape features regulate the spillover of beneficial insects between forest remnants and adjacent coffee plantations.
Medeiros et al. (2019a) collected bees, flower flies and predatory wasps in 16 coffee-producing landscapes to investigate how landscape structure affects alpha and beta diversity of beneficial insects. In each of the 16 landscapes, beneficial insects were collected in forest remnants and adjacent coffee plantations and all individuals collected in both habitats were aggregated to calculate alpha and beta diversity at landscape level. For predatory wasps the authors found that total species richness and abundance increased, and species loss decreased with increasing landscape forest cover. Here, we used the same dataset as Medeiros et al. (2019a) to assess the effects of landscape composition and configuration on the spillover of wasps between forest remnants and adjacent coffee plantations. We used incidence and abundance-based dissimilarity indices to estimate the inter-patch movement of wasp species and individuals and we assumed that high dissimilarity means less flow (and thus less spillover) between forest and coffee habitats.
We hypothesized that the dissimilarity of paired wasp communities between forest and coffee habitats decreases with increasing forest cover, landscape diversity and edge density in the surrounding landscape, whereas pesticide usage does not affect dissimilarity patterns. We expected that forest cover would increase wasp spillover between habitats by increasing the availability of unmanaged nesting sites, whereas high diversity of habitat types and edge environments would provide alternative and complementary feeding habitats. We also expected that the spillover of wasps between forest and coffee habitats is mainly regulated by landscape features that minimize the negative impact of pesticide usage in crop matrix. We provide important information on how landscape structure regulates the spillover of beneficial insects in forest remnants and adjacent crops. Such information is crucial for landscape planning and decision-making focused on promoting the synergy between biodiversity conservation and the provision of ecosystem services in coffee-producing landscapes.
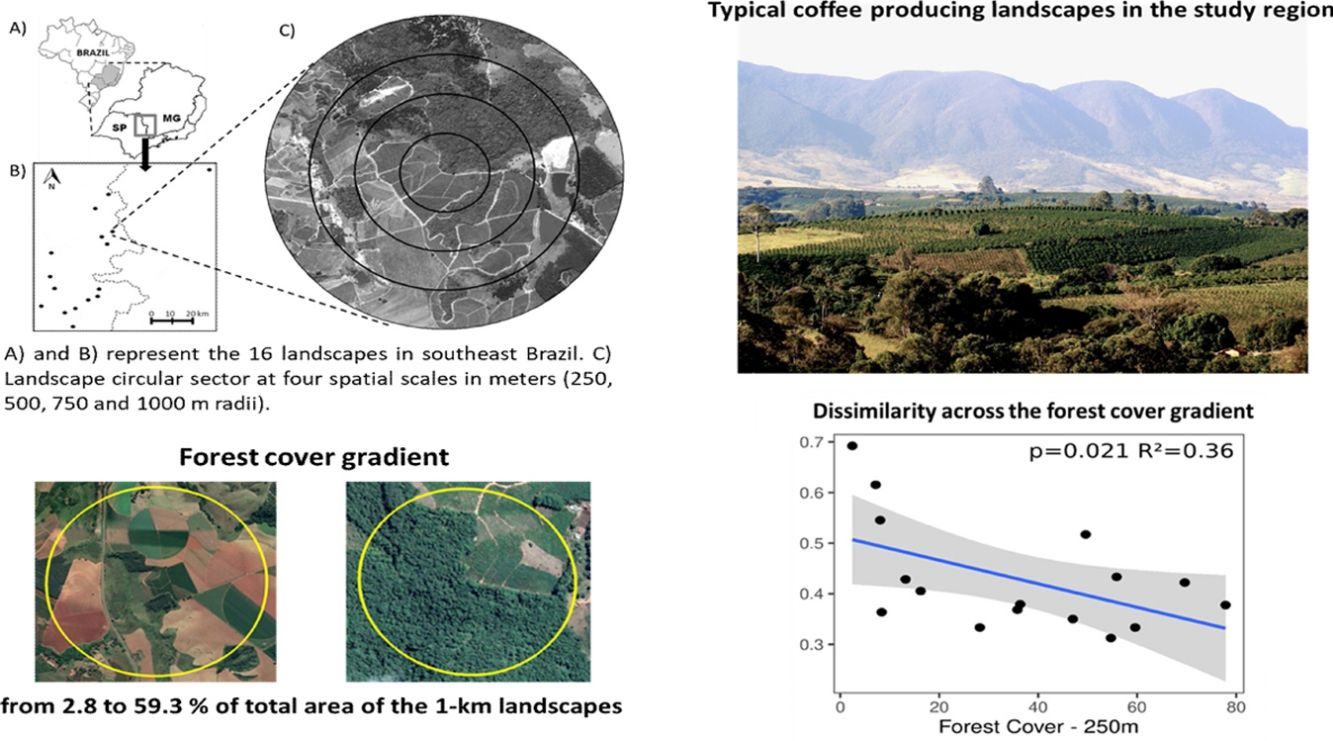
MethodsStudy areaThe study was conducted in 16 non-overlapping coffee producing landscapes near the border of the states of São Paulo and Minas Gerais in southeastern Brazil (Fig. 1A). The 16 landscapes compose a gradient of landscape composition and configuration from forested landscapes to crop dominated landscapes. The study region is an important coffee producing region in the country and it was responsible for 13% of the national production of Coffea arabica L. in 2018–2019 (Conab, 2019). The predominant vegetation in the study region is the seasonal semideciduous forest, an ecosystem type of the Atlantic Forest biome that was heavily converted into sun-coffee plantations, pastures and eucalyptus stands.
, the spatial arrangement of our 16 landscapes within the study region (B) and one of our 16 1km-landscapes (C). In (A), SP = Sao Paulo and MG = Minas Gerais states. In (C), the black rings represent the four circular spatial scales used in this study (250, 500. 750 and 1000 m radii) our forest remnants and adjacent coffee plantations are centrally located in each landscape within the 250 m radius. Yellow lines show linear transects where traps were set up in each habitat.")
The location of our study region in southeast Brazil (A), the spatial arrangement of our 16 landscapes within the study region (B) and one of our 16 1km-landscapes (C). In (A), SP = Sao Paulo and MG = Minas Gerais states. In (C), the black rings represent the four circular spatial scales used in this study (250, 500. 750 and 1000 m radii) our forest remnants and adjacent coffee plantations are centrally located in each landscape within the 250 m radius. Yellow lines show linear transects where traps were set up in each habitat.
Wasps (Vespidae: Polistinae and Eumeninae) were collected in forest remnants and in adjacent coffee monocultures. In each landscape, we selected one forest remnant and an adjacent coffee plantation centrally located in each of the 16 landscapes (Fig. 1B and C). In each forest and coffee habitat, we set up four traps – one Malaise trap (Townes model) and three baited traps – that were placed along a linear transect parallel to the edge with the paired coffee or forest habitat. The linear transects were placed at a fixed distance of 100 m from the target edge and at least 100 m away from any other edge. In each transect the four traps were 50 m apart. We used 2 l plastic bottles with four circular 4 cm diameter holes cut midway as baited traps that were individually filled with 300 ml of orange juice and 0.4 ml of propylene glycol. Baited traps were hung from branches at 1.3 m above the ground. Wasps were collected for seven consecutive days per month for 13 months from June 2016 to June 2017. Captured wasps were preserved in 70% ethanol and identified to the lowest taxonomic level possible using available identification keys (Supplementary material S1). We also performed monthly interviews with farmers to record the number of insecticides, herbicides and fungicides applications per month during the 13 months of wasp surveys in each of the 16 coffee plantations where wasps were collected. The cumulative number of pesticide applications over the 13 months of wasp surveys varied from 2 to 13 applications per coffee plantation.
Landscape variablesWe built land use maps using ArcGIS basemap imagery, Digital Globe satellites (1 m resolution) from 2013 to 2015. Satellite images were projected in UTM SAD 1969 at a 1:5000 spatial scale. We also performed extensive ground verification to improve the accuracy of land use maps. We mapped 10 land-use types (in alphabetical order): abandoned crops, abandoned pastures, arable field, built up area, coffee, eucalyptus, forest, pasture, water body and wetland. These land use types were used to calculate the following explanatory landscape variables: edge density (landscape configuration), and forest cover and landscape diversity (landscape composition) at four spatial scales (250, 500, 750, and 1000 m radius). These spatial scales are suitable for wasps in Brazilian agricultural landscapes (Medeiros et al., 2019a). Forest cover encompassed the total area of forest in the landscape transformed to percentage cover. Landscape diversity was calculated using the Shannon diversity index. For a full description of Shannon landscape diversity and edge density indices see McGarigal (2015). Landscape variables were calculated using ArcGIS 9.3 with Patch Analyst extension (Rempel et al., 2012). At a 1 km landscape scale, forest cover varied from 2.8 to 59.3%, landscape diversity from 1.05 to 1.86 and edge density from 127 to 246.
Statistical analysesWe used incidence- and abundance-based beta diversity partitioning approaches to estimate wasp spillover between forest and coffee habitats. The incidence-based dissimilarity approach consists of decomposing total beta diversity (Sorensen dissimilarity index) into two components: nested and turnover, which reflect species loss and species replacement, respectively (Baselga, 2010). The turnover component is represented by the Simpson dissimilarity index, whereas the nested component is the difference between Sorensen and Simpson indices (Baselga, 2010). Analogous to the incidence-based beta diversity, the abundance-based dissimilarity approach consists of decomposing the Bray-Curtis dissimilarity index into two components: abundance gradient and balanced variation, which represent the loss and replacement of individuals, respectively (Baselga, 2013). We focused on Sorensen and Bray-Curtis indices and their turnover and balanced variation components because these indices are directly associated with the inter-habitat spillover, whereas nested and abundance gradient are associated with loss of species and specimens, respectively. We assumed that high dissimilarity means that paired communities share fewer species (Sorensen and Simpson indices) and individuals (Bray-Curtis and its balanced variation) indicating no spillover between habitats.
To investigate the effects of landscape composition and configuration on wasp spillover we performed linear models using Sorensen and Bray-Curtis dissimilarity indices and their components in each habitat (forest and coffee) as response variables. For each response variable we generated 12 models using the three-landscape metrics (forest cover, landscape diversity and edge density) at four spatial scales (250, 500, 750 and 1000 m radius). Since pesticide usage is a well-known factor affecting non-target beneficial insects (Desneux et al., 2007), we included this variable in all 12 models to identify significant interactions between local pesticide usage and landscape variables at multiple spatial scales. We determined the scale of effect of each landscape metric by using the spatial scale with the highest coefficient of determination (Jackson and Fahrig, 2015). We considered that a given predictor affects wasp dissimilarity between habitats when this predictor fitted a significant model (p < 0.05) and the confidence interval of the slope did not include zero (Arnold, 2010). If the variable pesticide usage did not present significant interactions with landscape predictors (slope includes zero), we generated new models without pesticide usage. In the case of two or more predictors generating significant models, we used the corrected Akaike Information Criterion with the AIC difference from the best model (Δ AIC) to select the best model in the set of competing models (Burnham and Anderson, 2002).
All analyses were performed in the software R version 3.3.2 (R Core Team 2014). We used the presence-absence community matrix as input in the function “beta.pair” and abundance community matrix as input in the function “beta.pair.abund” both from the “betapart” package (Baselga and Orme, 2012) to calculate incidence and abundance based dissimilarities, respectively. We also used “bbmle” package (Bolker, 2010) for model selection and “ggplot2” package (Wickham, 2016) to build graphs.
ResultsWe collected a total of 9847 wasps (Vespidae: Polistinae and Eumeninae) classified into 75 species and 23 genera (Fig. 2 and supplementary material S1). Wasp species richness was higher in coffee plantations (68 species) than in forest remnants (43 species). Wasps were also more abundant in coffee plantations (6376 individuals) than in forest remnants (3471 individuals). The species Agelaia multipicta (Haliday, 1836) and Agelaia vicina (de Saussure, 1854) were dominant in both habitats and together encompassed 66% of the individuals collected in forest remnants and 40% in coffee plantations. Landscape explanatory variables and pesticide usage varied considerably along the 16 landscapes.
 in southeast Brazil. The letters C and F represent the paired coffee and forest habitats in each of the 16 sites/landscapes.")
All four dissimilarity indices (Sorensen, Simpson, Bray-Curtis and its balanced variation component) decreased with increasing forest cover at the finer scale (250 m - Fig. 3) suggesting spillover of wasps between forest and coffee habitats increased with increasing forest cover in the surrounding landscape. The explanatory variables landscape diversity, edge density and pesticide usage did not affect wasp dissimilarities between forest remnants and coffee plantations. The nested and abundance gradient components did not respond to any of the explanatory variables (Supplementary material S2).

Linear models representing the influence of landscape composition on spillover of wasps between forest remnants and adjacent coffee plantations as represented by the influence of forest cover on dissimilarity indices. Incidence-based indices are represented by Sorensen and Simpson dissimilarity indices, respectively, for A and B. Abundance-based indices are represented by Bray-Curtis index and its balanced variation component, respectively for C and D.
Wasp dissimilarity decreased with increasing forest cover in the surrounding landscape, suggesting that forest amount increases the spillover of wasps between forest remnants and adjacent coffee plantations. In contrast, wasp dissimilarity did not respond to landscape diversity or edge density.
Beneficial insects encompass many functional groups that perceive landscape composition and configuration in different ways. Some species rely on landscape complementation (Dunning et al., 1992) and benefit from landscape diversity and edge environments, while other species rely on habitat amount (Fahrig, 2013) and strongly dependent on a single habitat or few habitat types. Our findings suggest that forest cover (habitat amount) in the surrounding landscape is the main factor regulating wasp spillover between forest and coffee habitats at a local scale. All four dissimilarity indices decreased with increasing forest cover in the landscape resulting in high diversity and equability between forest and coffee habitats. This means that forested landscapes support more diverse wasp communities because of the high habitat amount that improves the accessibility of local communities to the regional species pool by increasing spillover opportunities and availability of nesting habitats.
For wasps, forest remnants are important sources of undisturbed nesting sites (Souza et al., 2014; Oliveira et al., 2017) and food resources (Sobek et al., 2009) that are crucial for wasp persistence in periods of scarcity in farmland. Nesting sites are particularly important for wasps, which are central foragers and move repeatedly among nesting and feeding habitats (Charnov, 1976; Richter, 2000). In our study system, we assumed that wasps use forest remnants as nesting habitats and coffee plantations as feeding habitats. We argue that coffee plantations cannot provide nesting habitats for wasps mainly due to nest removal that is a common practice among farmers to avoid sting accidents during crop management and harvesting operations. At the habitat level, coffee plantations supported higher species richness and abundance of wasps than in adjacent forest remnants. Such a pattern can be associated with high primary productivity of crops leading to a higher density of consumers in the crop matrix than in adjacent natural habitats (Rand et al., 2006; Frost et al., 2015). However, we highlight that wasp alpha diversity in both forest and coffee habitats increased with increasing forest cover revealing that crop dominated landscapes supported only subsets of species found in forested landscapes (Medeiros et al., 2019a).
The lack of effect of pesticide usage on wasp dissimilarity indices can be also associated with the rapid recolonization of crops promoted by the spillover from forest remnants (nesting habitats; Souza et al., 2010, 2014) that act as permanent sources of wasps to adjacent coffee plantations (Medeiros et al., 2019b). Moreover, landscape diversity and edge density did not affect wasp spillover suggesting that nesting sites are not available in other crop and non-crop habitats nor in edge environments.
The four dissimilarity indices (Sorensen, Bray-Curtis, Simpson, and Balanced variation) responded to forest cover at the smaller scale. This points out that most wasp species can spillover between habitats within landscapes at a radius of 250 m that is in accordance with the very few studies available from the Neotropical region suggesting that some wasp species have a dispersal range of up to 300 m from nesting sites (Santos et al., 2000; da Cruz et al., 2006, Ribeiro-Filho et al., 2008). However, we highlight that the alpha and beta diversity of wasps at the landscape level responded to forest cover up to 1000 m (Medeiros et al., 2019a). Therefore, the effects of forest cover on the landscape wasp species pool seems to occur at larger spatial scales.
Forest cover reduced species loss and increased diversity of wasps and provision of biological control services in the same 16 landscapes used in the present study (Medeiros et al., 2019a, 2019b). Forest cover is also important for other beneficial insects and their ecosystem services in coffee producing landscapes. In our study region, Saturni et al. (2016) found an increment of 28% in coffee fruit set that was associated with the presence of bees, which were positively correlated with forest amount in the surrounding landscape. Similar results were found for predatory ants, which decreased presence, infestation, and damage of a major coffee pest in forested landscapes, but not in structurally simple landscapes (Aristizábal and Metzger, 2019). The positive effects of forest remnants on beneficial insects and the provision of ecosystem services in adjacent coffee plantations were also observed in Central America (Roubik, 2002; Ricketts, 2004), southeast Asia (Pries et al., 2007; Krishnan et al., 2012) and Africa (Milligan et al., 2016; Berecha et al., 2015). Therefore, forest conservation and restoration initiatives should be incorporated in landscape planning and management to improve the spillover of beneficial insects and their ecosystem services in farmland creating a win-win scenario for biodiversity and coffee farmers.
Authors’ contributionsThe study was designed by H.R.M, C.A.R and M.C.R.; Data collections were performed by H.R.M.; Data analyses were performed by F.M. All authors actively contributed to writing and revising the manuscript.
Conflict of interestThe authors declare that there are no competing personal and financial interests.
We thank Dr. Yuri Campanholo Grandinete who identified all wasps collected in this study and Rufford Foundation that provided crucial financial support for fieldwork activities (reference project: 18799-1). We are grateful to the owners of private lands where the study sites are located. We also thank the editor and the reviewers who made valuable comments and suggestions that substantially improved the manuscript. HRM received a research grant from Brazilian Government Research Council (CNPq) (142147/2015-0/141932/2016-3) and a scholarship from Emerging Leaders of Americas Program (ELAP) supported by Canadian Government. JPM was supported by a FAPESP grant (Interface Project, N. 2013/23457-6) and by the Brazilian Science Council (CNPq; grants 306121/2016-6, and 305484/2017-6). MCR thanks to FAPESP (process #2013/50421-2; #2020/1779-5; #2021/08534-0), CNPq (processes #312045/2013-1; #312292/2016-3; #442147/2020-1) and PROCAD/CAPES (project # 88881.068425/2014-01) for their financial support.
The following are Supplementary data to this article:

