
The framework species method has been successfully used to restore tropical forests in different parts of the globe. However, establishing experimental plantations for selecting a set of appropriate framework species is extremely time and labor consuming. Here we tested if species performance during colonization of degraded areas could be used to select framework species for restoration programs. We made a survey of the vegetation naturally colonizing a lake margin impacted by bauxite tailings. We followed survival and growth of these individuals during two years and compared their performance with planted individuals of the same species on a restoration project developed nearby. We found no clear relation between species performance during natural colonization and performance when planted in the same substrate. Future studies evaluating performance in the nursery and experimental plantations are still essential for selecting the best framework species and will greatly contribute for restoration of highly diverse ecosystems.
Land use changes exert strong impacts on biodiversity worldwide (Geist and Lambin, 2002; Sodhi, 2008). This is particularly true for developing countries, which harbor most of the global biodiversity hotspots (Mittermeier et al., 2004). At the same time, their vigorous economic growth promotes accelerated rates of land conversion. Therefore, developing successful strategies for restoring highly diverse tropical ecosystems are urgently needed (Dias et al., 2012; Rodrigues et al., 2011). This is recognized by the Convention of Biological Diversity that has set as one of its 20 targets for 2020 to restore 15% of all degraded habitats in the planet (Mittermeier et al., 2010). However, high diversity poses an additional challenge for ecosystem restoration, as a large number of species needs to successfully establish for recovering the typical structure and function of the system.
The use of framework species might help overcoming this problem (Goosem and Tucker, 1995; Lamb et al., 1997). By planting a restricted number of native species, it is possible to re-establish a multi-layered canopy structure and restore nutrient and carbon cycles. This new basic forest framework will then attract wildlife which can bring seeds and slowly increase forest diversity and functionality. While this method has been successfully used to restore tropical forests in different parts of the globe (Knowles and Parrotta, 1995; Tucker and Murphy, 1997; Wydhayagarn et al., 2009), a critical step is the selection of the appropriate framework species. The selection of framework species usually follows a set of criteria, namely: (i) high survival and growth of sapling planted in the degraded area, (ii) easy propagation by seeds under nursery, (iii) crown architecture to shade out weeds or improve understory environmental conditions and (iv) resistance to specific field harsh conditions (e.g., drought, fire and flooding). The evaluation of these criteria is, however, extremely time and labor consuming as it requires experimental plantations to follow species performance in the field (Elliott et al., 2003).
Alternatively, we suggest that identifying species with high performance during initial colonization in degraded areas could give a first hint of a set of framework species. If species performances during passive and active restoration are correlated, studying natural colonization of degraded areas would provide a cheaper method for selecting framework species as compared to experimental plantations. To test this hypothesis, we studied the natural colonization on an area impacted by bauxite tailings in the Amazon. Our approach was three-fold: first, we made a vegetation survey to identify species colonizing the bauxite tailings; second, we followed survival and growth of the naturally recruited species during two years; and third, we compared the performance of individuals of the same species naturally colonizing the bauxite tailings and planted on a restoration project developed nearby.
Materials and methodsStudy siteOur study site comprised an area impacted by bauxite tailings at the margins of Lake Batata (1°30′S, 56°20′W), which is located in the district of Porto Trombetas, municipality of Oriximiná, state of Pará, in the Brazilian Amazon. This lake lies on the right bank of the Trombetas, a clear water river. Clear-water rivers typically have high transparency, low quantity of organic and inorganic suspended material, a pH from low to neutral, low nutrient availability, and are bordered by igapó vegetation (see Prance, 1979 for terminology regarding Amazonian flooded forests).
As a result from bauxite mining in the nearby terra-firme forest, tailings were continuously discharged into the Lake from 1979 until 1989. When discharge was halted, ca. 600ha of lake (ca. 30% of the area of the lake) and marginal igapó (flood-prone) vegetation were buried by bauxite tailings. The bauxite tailings consist of red, muddy clay composed basically of iron and aluminum oxides and differ from native igapó soil in having a marked compaction, decreased porosity and infiltration capacity and reduced microbial activity (Parrotta and Knowles, 1999). The study site is subjected to eight months of flooding per year. This severe flooding regime represents a major challenge for restoration due to its impact on trees survival and growth.
On the study site, two restoration strategies were adopted. Passive restoration, where plants were allowed to freely colonize the substrate, and active restoration, where saplings of native igapó species were introduced. The areas of passive and active restoration were contiguous and had no differences in substrate, flooding regime and were not subjected to any management except for the sapling introduction in the active restoration area.
Passive restorationTo record the natural colonization of the bauxite tailings, we set up a 1.12ha grid, consisting of 28 quadrats, each with an area of 20m×20m. In December 2003, we labeled and identified all plants within the grid, which were then measured for height and basal diameter. Diameter of each individual was measured at the closest height of soil level that did not show any stem deformity (e.g., root above-ground expansions). In the following two years, in December 2004 and 2005, all surviving plants were measured again, and newly germinated plants were labeled, identified, measured and added to the list. Growth was calculated as the absolute increase in basal diameter per year. Since we found out that all newly germinated plants sampled in the second and third years had a basal diameter equal or lower than 0.5cm, we classified all such plants, including those sampled in 2003, as regenerants. Plants with values higher than 0.5cm were classified as recruits.
We calculated the importance value (IV; Müller-Dombois and Ellenberg, 1974) for the recruited plants of each species for all years. This is a measure of species dominance, which is given by the sum of relative frequency, density and basal area. We considered the species that accounted for up to 75% of total IV to be the dominant species and those with IV smaller than 0.1 to be locally rare species.
Active restorationIn December 2003, we established a species introduction experiment in an area adjacent to the plant survey. Saplings were grown in a greenhouse with ad libitum water and nutrients. On the time of introduction in the field, saplings varied from 30 to 50cm depending on the species. Introduced plants were placed in holes of 0.15m of diameter, with 0.2m of depth, opened with 2m tall heavy wooden stakes. These plants received no fertilization or irrigation after planting. During the subsequent two years, saplings were controlled for survival and measured for height and basal diameter as described above.
Statistical analysesWe used Pearson correlation coefficient to test if species performance (survival and growth) during passive restoration was correlated to the performance during active restoration. Also, using paired t-test, we tested if species differ in survival and growth between passive and active restoration.
ResultsPassive restorationThe number of species found in this plant community increased from 15 in 2003 to 32 woody species (27 trees, 1 shrub and 4 vines) in 2005 (Table S1). This was mostly due to a very marked increase in the number of regenerants in 2004 in relation to 2003 (5-fold) and in 2005 in relation to 2004 (4-fold). However, the number of recruited species remained rather constant (between 12 and 13). This is apparently more related to slow growth of these new-coming individuals than to mortality rates (only 3.5% for recruits as a whole over the two study years).
These 32 species recorded in the 2005 survey belonged to ten different families, and the Fabaceae was particularly species-rich (7 species, i.e. ca. 21% of the species total) (Table S1). There were no changes in the dominance structure of the community over the years (Table S2), since the dominant species (those that concentrated 75% of the IV) were the same (Buchenavia, Genipa, Dalbergia, and vine sp. 1).
There was an increase (60.6%) in recruit canopy cover between 2003 and 2005 (Table S2), which resulted mainly from a 15.5% rate of seedling recruitment (see Table S1), a very low mortality rate (3.5%), and an overall positive growth rate, particularly for the dominant species (Table 1).
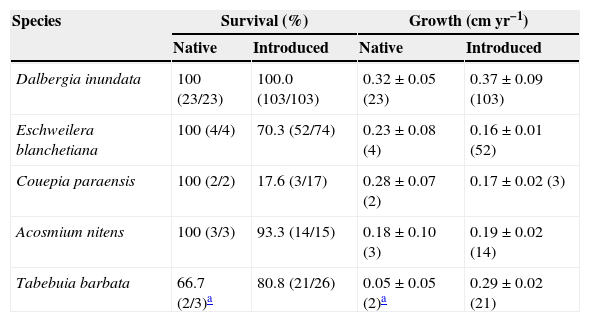
Survival and basal diameter growth [mean±standard error (n)] of introduced saplings of tree species on the bauxite tailings substrate as compared to natural regenerants over the study period (December 2003–December 2005). For this comparison we considered only introduced saplings and natural regenerants within the same range of diameter (0.2–1.5cm), since growth can vary with plant size.
| Species | Survival (%) | Growth (cmyr−1) | ||
|---|---|---|---|---|
| Native | Introduced | Native | Introduced | |
| Dalbergia inundata | 100 (23/23) | 100.0 (103/103) | 0.32±0.05 (23) | 0.37±0.09 (103) |
| Eschweilera blanchetiana | 100 (4/4) | 70.3 (52/74) | 0.23±0.08 (4) | 0.16±0.01 (52) |
| Couepia paraensis | 100 (2/2) | 17.6 (3/17) | 0.28±0.07 (2) | 0.17±0.02 (3) |
| Acosmium nitens | 100 (3/3) | 93.3 (14/15) | 0.18±0.10 (3) | 0.19±0.02 (14) |
| Tabebuia barbata | 66.7 (2/3)a | 80.8 (21/26) | 0.05±0.05 (2)a | 0.29±0.02 (21) |
However, only 2.12% of the area was covered by vegetation (canopy cover) after approximately ten years since this substrate consolidated, which is a clear indication that human intervention is needed.
Species performance in active vs. passive restorationFrom the species introduced in the experiment, five were also found naturally colonizing our study site (Table 1). There were no correlation on species survival (r=−0.14; p=0.82; df=3) and growth (r=0.04; p=0.94; df=3) between natural colonization and experimental introduction. There were no significant differences on growth (t=−0.36; p=0.74; df=4; paired t-test) and survival (t=1.24; p=0.28; df=4; paired t-test) between active and passive restoration.
DiscussionTo our knowledge, this is the first study testing the use of species performance during passive restoration for selecting framework species. Our results indicate that this is not always possible, as shown by the lack of relationship on species survival and growth between introduced and naturally recruiting plants. The search of fast and cheap alternative methods for selecting framework species is urgently needed, but our results indicate that plant surveys cannot replace experimental plantations for this purpose. Similar studies in different ecosystems would allow us to assess the generality of these findings.
Abundance during colonization of the bauxite tailings was also not a good indicator for performance during active restoration. If on the one hand, the abundant legumes Dalbergia and Acosmium performed well as introduced species, other abundant species either did not show good results after introduction (e.g., Couepia) or proved difficult to cultivate in the nursery (e.g., Buchenavia; data not shown). Moreover, some of the best results in terms of survival and growth in planted conditions were shown by Myrciaria (data not shown), which can be found in nearby non-impacted areas but was never found naturally recruiting on the bauxite tailings. This lack of a direct relationship of natural abundance with performance in nursery or after introduction has also been found for a Brazilian restinga, an oligotrophic habitat in the Brazilian coastal sandy plains (Zamith and Scarano, 2006).
Although we suggest a limited relevance of studying colonization of degraded areas for selecting framework species, such studies, together with vegetation surveys on pristine areas, are still important for building a first list of species to be tested experimentally. But germination tests and evaluation of performance in the nursery and experimental plantations are still essential for selecting the best framework species (Elliott et al., 2003; Kuaraksa and Elliott, 2013). This is especially true for ecosystems with given harsh conditions, such as flooding, drought or low nutrient availability. Also, the tropical region comprises many different ecosystem types which are still poorly studied. This combination of numerous ecosystem types with scarce information on their functioning makes it difficult to determine reference pristine systems, target species or ecosystem functions to be restored and lists of framework species for restoration programs. This ecosystem specificity highlights the urgent need to increase investments on restoration research on the tropics, where most of the biodiversity is concentrated (Aronson et al., 2010; Rodrigues et al., 2011). Future studies listing suitable framework species for different tropical ecosystems will greatly contribute to restoration of highly diverse ecosystems.
Conflicts of interestThe authors declare no conflicts of interest.
We thank Mineração Rio do Norte for funding and logistic support; and Brazilian Research (CNPq) and Education (CAPES) Councils for research grants. Dias ATC is supported by the Program Science without Borders (Bolsista CAPES/BRASIL).
The following are the Supplementary data to this article:

