





Human pressures such as hunting and habitat destruction can generate a deep fear in animals and this fear can influence their diel activity patterns and use of space. However, whether these behavioural responses to human pressure are consistent among key functional groups has been poorly studied. For example, while mammal species tend to become more nocturnal in areas with high human pressure, it is unclear if co-occurring birds display similar or opposite patterns. Here we used information from camera trapping (367 camera stations and 16,939 camera/days) along a gradient of human pressure in the Colombian Llanos to assess diel activity changes in understory birds and mammals. We found that diel activity significantly changed with higher human pressure for 45% of the birds (five species) and 36% of the mammals (five species) assessed, with four of five birds becoming more diurnal and all five mammals becoming more nocturnal. The average increase in nocturnality for the mammals was 11.3% while the average increase in diurnality for the birds was 7%. There was high variation in body size and dietary guild within impacted species, and only some were directly persecuted or hunted, suggesting that there are different pathways through which human pressure can affect vertebrates’ activity patterns. The contrasting behavioural responses to humans among vertebrate functional groups has significant repercussions for the fields of community ecology, including intraguild predation and competition, and should be a significant ecosystem-level conservation consideration.

Human impacts on nature have increased dramatically in the last 20 years, with over two thirds of global land area now facing moderate or intense human pressure (Williams et al., 2020). These pressures (e.g. hunting), along with related habitat changes have vastly altered wildlife communities through extirpations, extinctions, and changes to ecosystem functioning (Amir et al., 2022a; Dirzo et al., 2014; Haddad et al., 2015). These direct impacts are also accompanied by indirect impacts that change the behaviour of remaining wildlife, posing a significant threat to species fitness and interactions as well as overall biodiversity and ecosystem functioning (Gaynor et al., 2018; Sih et al., 2011; Zanette and Clinchy, 2019).
Human presence and habitat disturbance (together ‘human pressure’ hereafter) can generate a deep fear in animals and this “landscape of fear” can influence their diel activity patterns and use of space (Laundre et al., 2010; Zanette and Clinchy, 2019). While there are some studies showing little impact from human pressure on animal behaviour (Pardo et al., 2021), in general medium and large mammals tend to avoid humans by increasing their nocturnality (Gaynor et al., 2018; Mendes et al., 2020) or by reducing their home range closer to urban environments (O’Donnell and Delbarco-Trillo, 2020). Bird behavioural responses to humans, on the other hand, are less understood and may differ from those of mammals, especially in the tropics (Chace and Walsh, 2006; Zanette and Clinchy, 2019). A study on Chilean forest birds found that several species shifted their diel activity peaks from early morning in old-growth forests to noon in plantations, and to the afternoon in logged forests (Fontúrbel et al., 2021) and another study on Hawaiian forest birds showed increased diurnality in fragmented landscapes (Smetzer et al., 2022). Moreover, a global review of the impacts of hunting and degradation on mammal and bird populations found consistent declines in mammal abundance as degradation and hunting increased, while birds showed the opposite trend (Osuri et al., 2020). Another global review of hunting in tropical forests found that birds and mammals both showed significant declines in hunting-accessible areas, although 25% less for birds than for mammals (Benítez-López et al., 2017).
There are different potential reasons why the ecology of fear may manifest itself differently across taxa (Zanette and Clinchy, 2019). Fearful animals may perceive safety differently depending on their ability to hide versus escape, with most birds preferring higher daytime visibility and ease of escape via flight, while terrestrial mammals may often view nocturnal stealthy foraging as safer (Chace and Walsh, 2006; Zanette and Clinchy, 2019). Body size and diet are other factors that can influence species responses to human pressure (Dehaudt et al., 2022; Dunn et al., 2022; Hendry et al., 2023; Osuri et al., 2020). Species with larger body sizes are more susceptible to human pressures, particularly hunting (Benítez-López et al., 2017; Dirzo et al., 2014; Osuri et al., 2020), so these species could be more prone to change their activity patterns in areas with human presence (Benítez-López et al., 2017; Osuri et al., 2020). Habitat degradation and conversion can also drive changes in species behaviour by altering resource availability for some dietary guilds (Osuri et al., 2020). In particular, changes in diurnality patterns may be the consequence of altered foraging patterns due to changes in food availability in degraded habitats [e.g. due to logging, edges, fragmentation (Fontúrbel et al., 2021; Norris et al., 2010; Smetzer et al., 2022)].
Understanding behavioural responses of birds and mammals to human pressure is an important knowledge gap. Synchronous or opposing guild-level shifts among mammals and birds can shape consequences for food-web dynamics and ecosystem functioning (Gaynor et al., 2018; Zanette and Clinchy, 2019), and can also have repercussions for species interactions through changes in predatorial behaviour and increased competition between ecologically similar species (Amir et al., 2022b; Mendes et al., 2020; Zanette and Clinchy, 2019). Here, we investigate the impacts of human pressure on mammal and bird behaviour in Colombia, a megadiverse country that has seen a doubling of human pressure levels in the last 50 years (Correa Ayram et al., 2020). We used camera trapping along a gradient of human pressure in the Colombian Llanos region to measure changes in diel activity patterns of understory birds and mammals. We expected to find an increase in nocturnality in mammals and in diurnality for birds, a more marked shift in diel activity patterns for larger species or those more susceptible to hunting, and consistent responses from species with similar dietary characteristics.
MethodsSurveyTo detect possible shifts in the activity patterns of understory birds and mammals, we used a large dataset of camera-traps (Bushnell Core DS No Glow) to record the activity period of bird and mammal species in areas with different levels of human pressure. The study area is located in the Orinoco region in Colombia. The region is highly biodiverse but also under increasing human pressure (Correa Ayram et al., 2020). Sampling was done concomitantly and occurred from November 2020 to February of 2021 in three areas with different levels of human pressure (Fig. 1). The sampling areas are all located in mosaic landscapes, ranging from continuous forest to isolated forest patches near human settlements and roads, swamps and natural savannah, and pasture for livestock with scattered trees and bushes.
 In red the location of the study areas in Colombia. In grey the Llanos region. Location of the three sites where cameras were deployed in the study area: (b) Meta Foothills. (c) Casanare Foothills. (d) Tillavá River. Camera trap stations are depicted by purple circles. The level of human pressure in the study area is depicted using the Legacy-adjusted Human Footprint Index (HFI) for 2018. Red colours depict areas of high human pressure while blue colours areas of low human pressure.")
(a) In red the location of the study areas in Colombia. In grey the Llanos region. Location of the three sites where cameras were deployed in the study area: (b) Meta Foothills. (c) Casanare Foothills. (d) Tillavá River. Camera trap stations are depicted by purple circles. The level of human pressure in the study area is depicted using the Legacy-adjusted Human Footprint Index (HFI) for 2018. Red colours depict areas of high human pressure while blue colours areas of low human pressure.
We sampled a total of 367 camera stations along a gradient of Human Footprint Index-HFI- (Correa Ayram et al., 2020), including 186 locations in the Meta Foothills site (22-85 HFI), 138 in the Casanare Foothills site (22-91 HFI) and 43 in the Tillavá River site (8-45 HFI). Each station received one camera for a period between 16 and 96 days with a mean of 45.2 days. Each camera trap station was placed no less than one km away from any other camera. The camera-traps were placed around 50 cm above the ground, recording photos in response to the activation of a passive infrared sensor. All images were stored and processed in the Wildlife Insights platform and specialists verified the animal identifications (Diaz-Pulido et al., 2022a, 2022b, 2022c). We used 30 min as a minimum time allowed between two records for the same species at the same camera, as a safety measure to avoid repeated records of the same individual. This effort resulted on a total of 16,939 camera/days and 165 species recorded; 120 birds and 45 mammals (SI Table 1).
With the data obtained from camera-trapping, we classified the records in two groups based on the time of sunset and sunrise. Records taken when the sun was up in the sky (i.e. after sunrise and before sunset) were considered “day” records, while records taken after the sunset or before sunrise were considered “night” records. Since the exact timing of sunrise slightly varies along the study area according to latitude and date, we used each camera station's longitude, latitude, date and time to obtain the exact timing of sunrise and sunset from the U.S naval observatory solar calendar (U.S. Naval Observatory, 2022). The records for each species were then classified in day/night. By assuming each day/night record to be a Bernoulli trial, we treated the data from the camera trap stations as a binomial variable (Mendes et al., 2020), and evaluated the effect of human pressure on 11 bird and 14 mammal species, which had enough data across the study area to perform the analysis (Table 1).
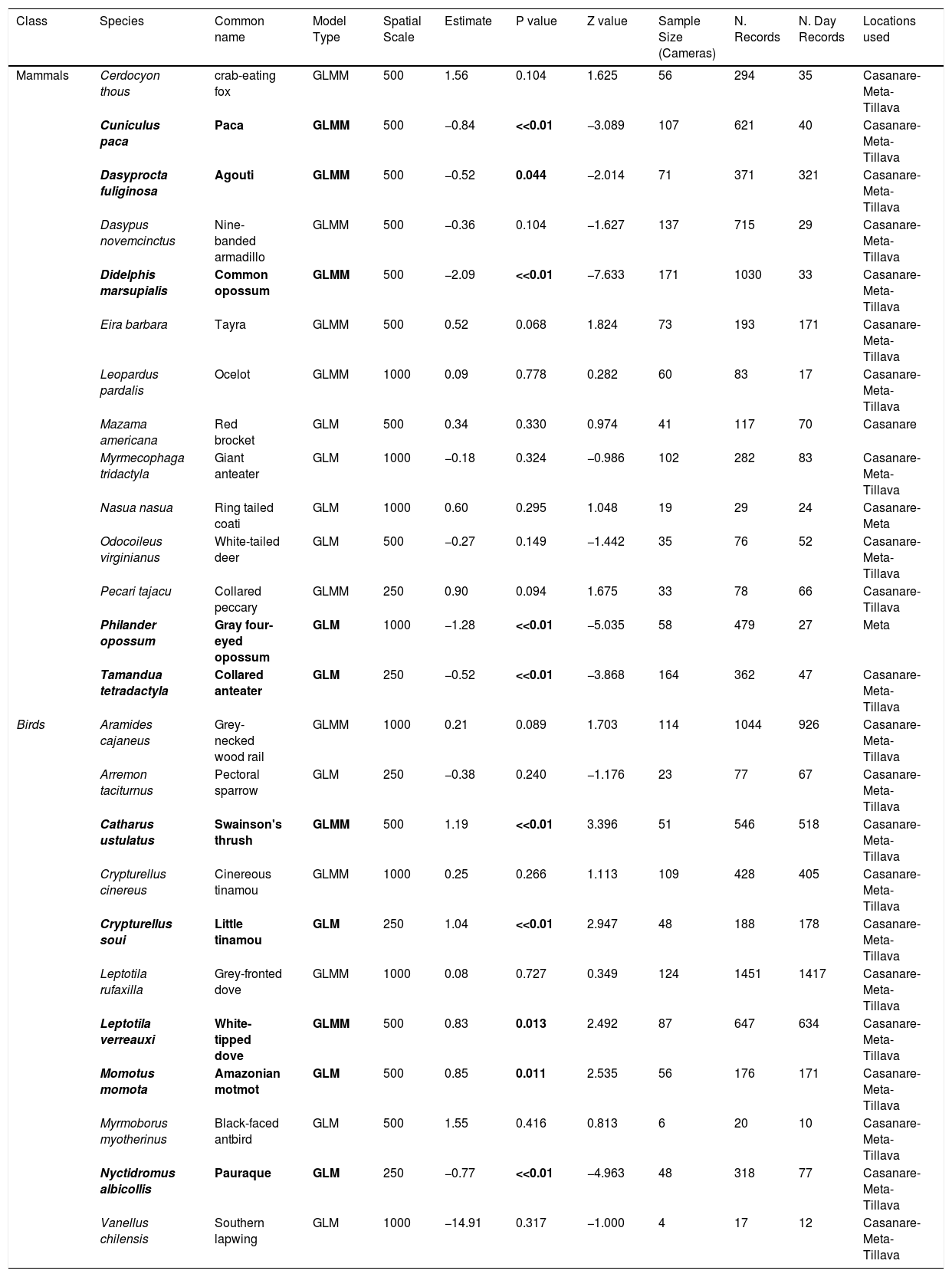
Main results of the generalized linear mixed models (GLMM) and general linear models (GLM) to evaluate the effect of human pressure on the diurnality of each understory mammal and bird species assessed. Significant results are in bold.
| Class | Species | Common name | Model Type | Spatial Scale | Estimate | P value | Z value | Sample Size (Cameras) | N. Records | N. Day Records | Locations used |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mammals | Cerdocyon thous | crab-eating fox | GLMM | 500 | 1.56 | 0.104 | 1.625 | 56 | 294 | 35 | Casanare-Meta-Tillava |
| Cuniculus paca | Paca | GLMM | 500 | −0.84 | <<0.01 | −3.089 | 107 | 621 | 40 | Casanare-Meta-Tillava | |
| Dasyprocta fuliginosa | Agouti | GLMM | 500 | −0.52 | 0.044 | −2.014 | 71 | 371 | 321 | Casanare-Meta-Tillava | |
| Dasypus novemcinctus | Nine-banded armadillo | GLMM | 500 | −0.36 | 0.104 | −1.627 | 137 | 715 | 29 | Casanare-Meta-Tillava | |
| Didelphis marsupialis | Common opossum | GLMM | 500 | −2.09 | <<0.01 | −7.633 | 171 | 1030 | 33 | Casanare-Meta-Tillava | |
| Eira barbara | Tayra | GLMM | 500 | 0.52 | 0.068 | 1.824 | 73 | 193 | 171 | Casanare-Meta-Tillava | |
| Leopardus pardalis | Ocelot | GLMM | 1000 | 0.09 | 0.778 | 0.282 | 60 | 83 | 17 | Casanare-Meta-Tillava | |
| Mazama americana | Red brocket | GLM | 500 | 0.34 | 0.330 | 0.974 | 41 | 117 | 70 | Casanare | |
| Myrmecophaga tridactyla | Giant anteater | GLM | 1000 | −0.18 | 0.324 | −0.986 | 102 | 282 | 83 | Casanare-Meta-Tillava | |
| Nasua nasua | Ring tailed coati | GLM | 1000 | 0.60 | 0.295 | 1.048 | 19 | 29 | 24 | Casanare-Meta | |
| Odocoileus virginianus | White-tailed deer | GLM | 500 | −0.27 | 0.149 | −1.442 | 35 | 76 | 52 | Casanare-Meta-Tillava | |
| Pecari tajacu | Collared peccary | GLMM | 250 | 0.90 | 0.094 | 1.675 | 33 | 78 | 66 | Casanare-Tillava | |
| Philander opossum | Gray four-eyed opossum | GLM | 1000 | −1.28 | <<0.01 | −5.035 | 58 | 479 | 27 | Meta | |
| Tamandua tetradactyla | Collared anteater | GLM | 250 | −0.52 | <<0.01 | −3.868 | 164 | 362 | 47 | Casanare-Meta-Tillava | |
| Birds | Aramides cajaneus | Grey-necked wood rail | GLMM | 1000 | 0.21 | 0.089 | 1.703 | 114 | 1044 | 926 | Casanare-Meta-Tillava |
| Arremon taciturnus | Pectoral sparrow | GLM | 250 | −0.38 | 0.240 | −1.176 | 23 | 77 | 67 | Casanare-Meta-Tillava | |
| Catharus ustulatus | Swainson's thrush | GLMM | 500 | 1.19 | <<0.01 | 3.396 | 51 | 546 | 518 | Casanare-Meta-Tillava | |
| Crypturellus cinereus | Cinereous tinamou | GLMM | 1000 | 0.25 | 0.266 | 1.113 | 109 | 428 | 405 | Casanare-Meta-Tillava | |
| Crypturellus soui | Little tinamou | GLM | 250 | 1.04 | <<0.01 | 2.947 | 48 | 188 | 178 | Casanare-Meta-Tillava | |
| Leptotila rufaxilla | Grey-fronted dove | GLMM | 1000 | 0.08 | 0.727 | 0.349 | 124 | 1451 | 1417 | Casanare-Meta-Tillava | |
| Leptotila verreauxi | White-tipped dove | GLMM | 500 | 0.83 | 0.013 | 2.492 | 87 | 647 | 634 | Casanare-Meta-Tillava | |
| Momotus momota | Amazonian motmot | GLM | 500 | 0.85 | 0.011 | 2.535 | 56 | 176 | 171 | Casanare-Meta-Tillava | |
| Myrmoborus myotherinus | Black-faced antbird | GLM | 500 | 1.55 | 0.416 | 0.813 | 6 | 20 | 10 | Casanare-Meta-Tillava | |
| Nyctidromus albicollis | Pauraque | GLM | 250 | −0.77 | <<0.01 | −4.963 | 48 | 318 | 77 | Casanare-Meta-Tillava | |
| Vanellus chilensis | Southern lapwing | GLM | 1000 | −14.91 | 0.317 | −1.000 | 4 | 17 | 12 | Casanare-Meta-Tillava |
We used the recently developed Colombian Legacy-adjusted Human Footprint Index (HFI) for 2018, which is the most recent product of its kind for Colombia. It is temporarily consistent, and has a higher spatial resolution (300 m) than any other national (Etter et al., 2011) or global human footprint map (Venter et al., 2016; Williams et al., 2020). This index measures the level of human pressure on ecosystems and provides a baseline for monitoring human pressure on biodiversity. It was generated by the Humboldt institute (Correa Ayram et al., 2019) using the same methodology as the one developed for the year 2015 by Correa Ayram et al. (2020). The HFI is estimated using seven primary variables: land use type, rural population density, distance to roads, distance to settlements, fragmentation of natural vegetation, the biomass relative to natural potential, and time of intervention on ecosystems in years. All seven primary variables were collated for the year 2018 and re-scaled between 0 and 5 to reflect their relative contribution to human impact and transformation of ecosystems. Values of 0 indicate a null contribution, while values of 5 indicated a very high contribution. To generate a final index, the variables were added and the index was normalized between 0 and 100. See Correa Ayram et al. (2020) for a complete description of the variables and the methodology. The HFI for each camera station was recorded.
Data analysisWe used generalized linear mixed models (GLMM) to evaluate the effect of human pressure on understory birds’ and mammals’ diel activity patterns, using the day/night records for each species as a binomial response variable, the HFI as the explanatory variable and the site as a random variable. When the species was detected in a single site or the sample size at a given site precluded the usage of a random variable (e.g., causing singular fit), we used generalized linear models (GLM) instead of GLMM. Species with less than 5 captures in either the day or night were also excluded from the analysis as the number of observations for these species was insufficient to allow for statistical comparison. To determine the best spatial scale for each species, four logistic models were created using the average HFI from buffers with radiuses of 250, 500, 750 and 1000 m around each camera trap station. The results at each spatial scale were compared using Akaike Information Criterion corrected for small sample sizes (AICc) (Burnham and Anderson, 2018), and for each species the model with smallest AICc value was used for the logistic regression. Since the model selection was used to select the best spatial scale for each species, but not the overall model performance, a null model was not required (Mendes et al., 2020). For the species which presented responses to human pressure, we also calculated their activity shift, here described as the difference in the model predicted diurnality between the most preserved and most disturbed areas in which the species were recorded. Finally, the activity period of all species was also plotted for visual inspection (Fig. 2), and circular metrics of diel activity, such as mean direction and Rho (i.e., a measurement of circular mean and dispersion, respectively) were calculated and reported (SI Table 2). The species’ diet, body size, activity pattern (diurnal, nocturnal or cathemeral) and whether it was a game species or not was determined based on both local expert knowledge and literature (Emmons and Feer, 1997; Hilty and Brown, 2001; Myers et al., 2022; Ocampo et al., 2021). All analysis were performed in R version 4.2.1 (R Core Team, 2022), using the packages “lme4” (Bates et al., 2015), “circular” (Agostinelli and Lund, 2022), and ggplot2 (Wickham, 2016).
 five understory mammals and (b) five understory bird species in three areas in the Colombian Llanos. White bars represent cameras deployed in more preserved sited, and dark grey bars represent cameras deployed in more disturbed areas. Light grey is the overlap in the circadian activity between the more preserved and more disturbed camara sites. For this figure, the location where a camera was deployed was considered disturbed when it presented a Human Footprint index (HFI) above the mean, considering only the sited where cameras were deployed that had recordings of the species. The species depicted are the ones that presented statistically significant changes in diurnality. Mammal and bird silhouettes were obtained from the PhyloPic public domain database (http://phylopic.org/).")
Circadian activity of (a) five understory mammals and (b) five understory bird species in three areas in the Colombian Llanos. White bars represent cameras deployed in more preserved sited, and dark grey bars represent cameras deployed in more disturbed areas. Light grey is the overlap in the circadian activity between the more preserved and more disturbed camara sites. For this figure, the location where a camera was deployed was considered disturbed when it presented a Human Footprint index (HFI) above the mean, considering only the sited where cameras were deployed that had recordings of the species. The species depicted are the ones that presented statistically significant changes in diurnality. Mammal and bird silhouettes were obtained from the PhyloPic public domain database (http://phylopic.org/).
We found evidence of significant activity shifts in response to human pressure for five of the 11 species of understory birds (45%) and for five of 14 species of the understory mammals (36%) that had enough records to be assessed. For mammals, all five species became more nocturnal in disturbed areas (Table 1, Fig. 3a & 4a), while in birds four became more diurnal while one became more nocturnal (Table 1, Fig. 3b & 4b). The average increase in nocturnality for mammals was 11.3% while the average increase in diurnality for birds was 7%. While these shifts are considerable, the statements “becoming more nocturnal” or “becoming more diurnal” do not mean that a species shifted its peak of activity from a period to another. Instead, it means that although the species main activity is still occurring during its natural activity period (i.e., night for nocturnal species and day for diurnal species), the proportion of day/night records has changed.
 five understory mammals and (b) five understory bird species in response to human pressure in three sites in the Colombian Llanos, from the most preserved sites (black point) to the most disturbed sites (arrow point) in which each species was recorded. Arrows to the right represent shifts toward diurnality, while arrows to the left represent shifts toward nocturnality. The species depicted are the ones that presented statistically significant changes in diurnality. Mammal and bird silhouettes were obtained from the PhyloPic public domain database (http://phylopic.org/).")
Shifts in diurnality of (a) five understory mammals and (b) five understory bird species in response to human pressure in three sites in the Colombian Llanos, from the most preserved sites (black point) to the most disturbed sites (arrow point) in which each species was recorded. Arrows to the right represent shifts toward diurnality, while arrows to the left represent shifts toward nocturnality. The species depicted are the ones that presented statistically significant changes in diurnality. Mammal and bird silhouettes were obtained from the PhyloPic public domain database (http://phylopic.org/).
 five understory mammals and (b) five understory bird species with increases in human pressure. We used generalized linear mixed models (GLMM) to evaluate the effect of human pressure on understory birds’ and mammals’ diel activity patterns, with the day/night records for each species as a binomial response variable and the Legacy-adjusted Human Footprint Index (HFI) for 2018 (Correa Ayram et al., 2019) as the predictor (x-axis). When the species was detected in a single site or the low sample size at a given site caused singular fit problems, we used generalized linear models (GLM) instead. The species depicted are the ones that presented statistically significant changes in diurnality. The blue circles are the independent day or night captures and the doted lines in the GLM show the 95% confidence intervals. Mammal and bird silhouettes were obtained from the PhyloPic public domain database (http://phylopic.org/).")
Changes in diurnal activity for (a) five understory mammals and (b) five understory bird species with increases in human pressure. We used generalized linear mixed models (GLMM) to evaluate the effect of human pressure on understory birds’ and mammals’ diel activity patterns, with the day/night records for each species as a binomial response variable and the Legacy-adjusted Human Footprint Index (HFI) for 2018 (Correa Ayram et al., 2019) as the predictor (x-axis). When the species was detected in a single site or the low sample size at a given site caused singular fit problems, we used generalized linear models (GLM) instead. The species depicted are the ones that presented statistically significant changes in diurnality. The blue circles are the independent day or night captures and the doted lines in the GLM show the 95% confidence intervals. Mammal and bird silhouettes were obtained from the PhyloPic public domain database (http://phylopic.org/).
The four understory bird species which responded to human pressure by becoming more diurnal were the swainson’s thrush (Catharus ustulatus), the little tinamou (Crypturellus soui), the white-tipped dove (Leptotila verreauxi) and the amazonian motmot (Momotus momota) (Table 1). None of the species weigh more than a kilogram, all of them have omnivorous diets and one is opportunistically hunted (the little tinamou; SI Table 2). The pauraque (Nyctidromus albicollis), which weighs less than a kilogram, is an insectivorous species and is considered crepuscular, was the only bird that responded to human pressure by becoming more nocturnal. The eight other bird species did not present statistically significant activity shifts in response to human pressure.
The five mammal species which responded to human pressure by becoming more nocturnal were the paca (Cuniculus paca), the agouti (Dasyprocta fuliginosa), the common opossum (Didelphis marsupialis), the grey four-eyed opossum (Philander opossum) and the collared anteater (Tamandua tetradactyla) (Table 1). These species’ body weight ranged from less than 1 kg–13 kg. Two of the species have an herbivorous diet (the agouti and the paca), two have an omnivorous diet (the common opossum and the gray four-eyed opossum), and one is mainly insectivorous (the collared anteater). Three of these species - the paca, the aguti and the common opossum - are hunted (SI Table 2). The other nine species did not present statistically significant activity shifts in response to human pressure.
DiscussionWe found evidence of significant activity shifts in response to human pressure for 45% of the understory bird species assessed, with four of five shifts being towards more diurnal activity patterns. This is one of the first studies to our knowledge that reports such a behavioural change for tropical understory birds, and our findings align with emerging evidence of increased diurnality patterns for birds in disturbed subtropical landscapes (Fontúrbel et al., 2021; Smetzer et al., 2022).
None of the bird species that showed a change in activity patterns had a body mass higher than 500 g, and only one was opportunistically persecuted (the little tinamou). This suggests that the increases in diurnality for these bird species may not be strongly associated with direct human pressures like hunting, and instead may be more related to habitat disturbance. This may be because degraded landscapes have a higher variation in microclimate and increased reach of sunlight to the understory, which can affect birds circadian clocks and activity patterns (Fontúrbel et al., 2021; Lehmann et al., 2012). Additionally, degraded landscapes tend to have altered resource availability, including lower insect abundance (Vergara et al., 2021) and decreased fruit production (Jansen et al., 2021), which can result in increased daytime foraging in diurnal bird species, specially at times with better sunlight, to compensate for decreased food availability (Fontúrbel et al., 2021; Smetzer et al., 2022). Increased diurnality could also be associated to the increased nocturnality of certain mammal predators (or other animals, e.g. snakes) not investigated in this study. The pauraque was the only bird species that showed a significant increase in nocturnality in landscapes with high human pressure. This species has activity peaks at dusk and dawn, when natural light intensity is low, meaning light pollution could affect this species as has been reported for other nightjar species (Sierro and Erhardt, 2019) and mammals (Mendes et al., 2020).
For mammals we found a divergent behavioural response to human pressure from that of birds, with all significantly impacted species becoming more nocturnal. These results align with other studies that show increased nocturnality for mammals in areas with high human pressure (Gaynor et al., 2018; Mendes et al., 2020). One of the factors that can influence mammal diel activity is hunting, as three of the five species with significant activity shifts are hunted or persecuted. Species with diel activity change varied in body size from less than 1 kg–13 kg. These suggest that the increases in nocturnality may not be more prevalent in larger mammal species in the study area.
Our results show that human pressure goes beyond impacting species richness and abundance and can also have a marked impact in the activity pattern of multiple vertebrate groups. Bird species had a general shift towards diurnality, while mammal species towards nocturnality, revealing that different vertebrate groups can react distinctly to increased human presence and activity. Changes in activity patterns can increase activity overlap with certain predators and ecologically similar species. This could increase predation risk and competition for resources (Reilly et al., 2017). On the other hand, predators can be affected by the decrease of prey availability due to shifts on the preys daily activity pattern (Ordiz et al., 2017; Reilly et al., 2017). Several of the understory species that presented significant activity shifts in our study area are prey of other vertebrates like ocelots and pumas. These shifts in activity patterns can potentially affect the predator’s fitness and survival.
There is a need for more research on the impact of human pressure on vertebrates’ behaviour and the causes of such shifts, as they could result in marked changes away from natural patterns, with consequences for fitness, population persistence, community interactions, and evolution (Chace and Walsh, 2006; Gaynor et al., 2018), as well as for conservation planning (Caro, 2016; Gaynor et al., 2018). A better understanding of these interactions would help identify which species are particularly susceptible to behavioural changes, which human activities are the most detrimental and at which times of the day this overlap is happening. This information can be then incorporated into conservation planning, for example through the implementation of restrictions to certain human activities during times of the day when species of conservation concern are most active or when the likelihood of negative human-wildlife encounters is highest (Gaynor et al., 2018).
The use of other types of surveying techniques including sound recordings, transects and field sampling will be important to assess the potential changes in diel activity patterns for mammals and bird species that prefer to use the canopy or midstory levels. The availability of camera trap information is increasing exponentially, but usually the only information reported is in relation to species’ community composition and population densities (Antunes et al., 2022). Our analysis highlights the fact that impacts of human pressure on behavioural characteristics may vary between species, individuals and sites, and that this variation can have impacts on the ecology and conservation of these species. Future studies should focus on the impact of human pressure on the behavioural patterns of vertebrates (Gaynor et al., 2018; Sih et al., 2011), especially for birds and other groups for which information in this regard is scarce (Chace and Walsh, 2006; Fontúrbel et al., 2021; Smetzer et al., 2022) and for biodiverse regions that are poorly surveyed.
Declaration of interestsThe authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Authors contributionsP.J.N and B.G.V conceived the idea; P.J.N and C.M designed the study; A.D.P and L.H.R collected the data; P.J.N and C.M performed the analysis; P.J.N wrote the original draft of the manuscript; All authors contributed comments and edits to the final paper.
We would like to acknowledge C. Rojano, L. Miranda y R. Ávila (Fundación Cunaguaro), A. Lopera, F. Hernández, R. Fernández y R. Rodríguez (Corporación Gaica), Á. E. Cáceres-Gómez, N. Ferro-Muñoz. F. Cáceres-Gómez (Biotica Consultores) for the field work, and S. Pérez for the revision of the bird species lists. We thank Ecopetrol S.A. and Instituto de Investigación de Recursos Biológicos Alexander Von Humboldt for financing the study within the framework of the FIBRAS Agreement.
The following is Supplementary data to this article:

