





Habitat suitability for mammal species in grassland/forest ecotones may be affected by changes in abiotic conditions (e.g. light incidence), grazing and burning disturbances, and woody encroachment. We evaluate models addressing the role of such factors on structuring non-volant small mammal assemblages considering (1) only disturbed and (2) all ecotones (disturbed and undisturbed). A complete model (i.e., abiotic gradients, disturbances and woody encroachment) was the most plausible for abundance considering all ecotones, and for niche similarity considering both all and only disturbed ecotones. Niche similarity increased with distance from hydric resources, and abundance with increasing vegetation height. Further, disturbed habitats harbored simplified species assemblages. Habitat selection was detected in all ecotones due to the occurrence of habitat-specialist species on undisturbed sites. We did not find an exclusive influence of woody encroachment on mammal diversity. Patterns described here are relevant for management of productive lands and for biodiversity conservation.
Habitat suitability is a pre-requisite for species occurrence. Selection of specific habitats – triggered by different niche requirements of species – generate non-random patterns of small mammal distribution across gradients (Kingston and Morris, 2000), which might influence the structure and ecological roles performed by mammal assemblages. Non-random patterns of species distribution along ecological gradients (as grassland–forest ecotones) may be an effect of changes on vegetation structure along edges, which alter edge permeability to abiotic factors (e.g. light, temperature), flux nutrients and individuals, and also alters resource distribution, availability and acquisition by species (Ries et al., 2004). Edge effects alter assemblage's diversity and lead non-volant small mammals to select specific microhabitats (Kingston and Morris, 2000; Wilson et al., 2010). Besides edge effects, moisture gradients seem related to the presence of refuges areas, which makes habitat colonization by small mammal populations more probable (Milstead et al., 2007). Changes in abiotic conditions along ecotones suggest a strong selection of preferable habitats by small mammals (Wilson et al., 2010). However, diversity patterns across grassland-forest habitats may be generated by processes that alter vegetation heterogeneity – e.g. grazing, woody encroachment – rather than abiotic gradients (Pedo et al., 2010; Sponchiado et al., 2012).
Ecological disturbances such as woody encroachment, burning and/or cattle grazing/trampling deeply alter the structure of grassland and forest habitats, and likely shape the spatial and temporal dynamics of ecotones in southern Brazil (Müller et al., 2012). Woody encroachment over grasslands is a natural process caused by higher precipitation and humidity during the Late Pleistocene-Holocene (4000 years-today) compared to Middle Pleistocene (≈10,000 years) (Behling et al., 2004). Warm climate favors forest expansion and has created, in some regions of southern Brazil, vegetation mosaics of grasslands interspersed with forests (Behling et al., 2004). Woody encroachment is mainly observed where disturbances such as fire and grazing have been suppressed (Müller et al., 2012). Forest expansion is considered a global threat to biodiversity of grassy biomes (Bond and Parr, 2010; Veldman et al., 2015) due to species turnover mediated by increases in the frequency of shrubs and trees on grasslands, allowing colonization of forest flora/fauna. In southern Brazil, management to prevent woody encroachment usually includes burning, grazing and mowing (Pillar and Velez, 2010). These disturbances affect the heterogeneity and suitability of forest and grassland habitats, and offer differential conditions for small mammal colonization, reproduction and food acquisition (Dunstan and Fox, 1996; Fox et al., 2003).
Woody encroachment and light disturbance also promote local heterogeneity by adding vegetation strata and increasing habitat complexity (Fox et al., 2003). Concerns about the management of grassland and forest habitats with disturbances – generally applied throughout landscapes – have arisen recently for southern Brazilian grasslands – Campos Sulinos (Luza et al., 2014) and for other regions worldwide (Andersen et al., 2012), because sites where disturbance are absent or at low levels (i.e., high vegetation, deep litter layer, presence of tree/shrub strata) are prefered by small mammals (Pedo et al., 2010; Sponchiado et al., 2012). Rodents and marsupials are the richest mammal orders in Campos Sulinos – together with Carnivora (Luza et al., 2015b). Ecological roles performed by small mammals include seed removal and dispersal, predation of seedlings, seeds, fruits and arthropods (Iob and Vieira, 2008), biological resistance against invasive plants (Muschetto et al., 2015), nutrient cycling, bioturbation, and as vectors of human diseases (Medan et al., 2011). Improper management may cause species losses and environmental desequilibria, which may alter both assemblage diversity and functional roles performed by mammals. Thus, grassland–forest ecotones are usefull to contrast hypothesis on the effect of abiotic gradients from hypothesis that depict different sources and levels of disturbances that alter habitats and the structure of small mammal assemblages.
The study aimed to identify processes that affect the diversity of non-volant small mammal assemblages from grassland–forest ecotones under different management regimes. We used species composition, richness, total abundance and niche similarity between co-occurring species as assemblage descriptors, and then built models that embraced: (1) abiotic gradients, (2) grazing/burning, and (3) woody encroachment. For species richness and for specialist species (differentiating between forest and grassland habitats) we expected a strong effect of abiotic gradients (mainly canopy openness). We expected a negative effect of grazing/burning for abundance, but a positive effect to generalist species and to niche similarity. We expected a positive effect of woody encroachment for forest mammals. We also built models disregarding ecotones where grazing/burning were absent for a long time (22 years), and expected a positive effect of grazing/burning on assemblage structure, due to higher frequency of mammals adapted to disturbances.
Materials and methodsStudy area and sampling of non-volant small mammalsThe study area comprised nine sites at Campos Sulinos (Fig. 1A), distributed along several grasslands and forest physiognomies in southern Brazil. We essentially considered ecotones under different intensities of burning and/or grazing disturbances, and where forest expansion occurs by the presence of nurse plants and forest patches within grasslands (Duarte et al., 2006).
 study region and sampled sites from Luza et al. (2015a), where: 1, 4, 7=Brazilian Highland Grasslands; 2, 5, 8=Serra do Sudeste; 3, 6, 9=Campanha (the numeric order represent the sampling order); (B) distribution of grids on sites; and (C) grid dimensions, where T: transect and P: point on Forest and Grassland habitats, from Luza et al. (2015a). Site Image (2) from Google Earth® (2016).")
Sampling scheme showing: (A) study region and sampled sites from Luza et al. (2015a), where: 1, 4, 7=Brazilian Highland Grasslands; 2, 5, 8=Serra do Sudeste; 3, 6, 9=Campanha (the numeric order represent the sampling order); (B) distribution of grids on sites; and (C) grid dimensions, where T: transect and P: point on Forest and Grassland habitats, from Luza et al. (2015a). Site Image (2) from Google Earth® (2016).
Captures took place from October through November 2011, January through April and September and December 2012, and February 2013. To evaluate a wide range of habitat characteristics, we regionally distributed our surveys (mean distance between sites=326.48±153.28km; longest distance=553km). In each of the nine sites, we established two 140×140m grids (Fig. 1B) at least 1km distant from each other (total 18 grids) and sampled each site during ten days (five days for each grid); eight transects 20m apart (four at forest and four at grassland) disposed along the interface between grassland and forest composed the grid (Fig. 1C). We employed as a basic sampling unit a transect composed of eight capture points – a point contained one Sherman (25×8×9cm) and one Tomahawk trap (45×17.5×15cm). We reset traps in the morning and afternoon, totaling an effort of 1280traps/grid. We measured, marked with small cuts on the ear, and released animals at the same capture point; identification of specimens at species level was performed using DNA sequencing (details in Luza et al., 2015a).
Habitat structureWe evaluated variables in each capture point, and then averaged at the transect scale. Abiotic gradients were characterized by distance from the nearest hydric resource (streams, marshes, rivers – in meters) and canopy openness (%). Litter depth (cm), height of the herbaceous layer (cm) and tree density – amount of tree branches and leaves touching a 1.5m height pole at 10-cm radius (cm) – described the effect of disturbances, because grazing/burning alters the expected relationship between these variables and grassland-forest gradient. The effect of woody encroachment was addressed by the proportion of points – from total capture points on each transect (points on patches/eight) – within large (area >2000m2) and small forest patches (area<2000m2), which form patches with some canopy and that are disconnected from continuous forest (Duarte et al., 2006). Spearman correlation between variables not exceeded 0.5 (Appendix A1a).
Niche similarityTo measure niche similarity, we used traits previously employed to highlight assembly patterns on small mammal assemblages (Luza et al., 2015a) by their relevance for habitat exploitation, resource acquisition and ecosystem processes. Traits differentiate walkers from jumpers (posterior foot length), fossorial from terrestrial (claw and ear size), and terrestrial from arboreal species (body/tail length ratio; Table A1). We also used male home range size to characterize dispersal capability – unsuitable habitats support only non-resident species (large home ranges/high dispersal ability). As sampled small mammals comprised mainly insectivore–omnivore species (≈ half of species – Table A1), we used a binary trait describing diet (1 – insectivore–omnivore; 0 – others). A binary trait also characterized species with morphologic adaptations for aquatic life (Nectomys squamipes, Scapteromys meridionalis).
Data analysisWe associated mammal diversity to environment in transects distributed across several sites. Temporally distributed samples used to increase spatial representativeness may cause a large temporal effect on assemblage structure and hide effects of interest. We tested these effects through multivariate analyses of variance. No temporal effect arose; instead, we registered a large variation in assemblage structure at the regional scale (Appendix A2). Furthermore, we founded absence or weak Mantel correlation between spatial distances and assemblage structure dissimilarity among transects (Appendix A3).
We maintained transects without records assuming absence as an effect of habitat quality (except for niche similarity – zero means that coexistent species have very dissimilar niches). Species richness was considered as the number of taxa in each sampling transect standardized by the net sampling effort of each grid (number of taxa/number of trapsavailable–number of trapsnon-available). Sampling efficiency was evaluated by Mao Tau (observed richness), Jackknife 1 and Chao 1 (and 95% confidence intervals – IC 95%) in software EstimateS v.8. Abundance was the total number of individuals captured in each transect. Because of a high correlation between species richness and abundance (r=0.9, p=0.001), richness was regarded as the regression residuals from the richness–abundance relationship.
The composition matrix containing Hellinger-transformed abundance within each sampling transect was submitted to Principal Coordinate Analysis (PCoA, on software Multiv v.2.95) in order to extract main axes of variation in species composition (>10% of total variation [see Appendix A4]). Niche similarity between co-occurring species was considered as the difference between Gini-Simpson index and Rao's Quadratic Entropy (Pillar et al., 2013), which expresses how much functional diversity deviates from species diversity (low niche similarity occurs when Rao's entropy approaches from Gini-Simpson diversity [i.e., when co-occurring species respond differentially to environment]).
We simultaneously compete models characterizing gradients of conditions and resources, burning/grazing effect, woody encroachment, multiple factors (complete model) and none factor (null model), for species composition, richness, abundance and niche similarity. We employed generalized linear mixed models (GLMM) in order to deal with large spatial variability in assemblage diversity and with the fact that grassland and forest differ in some habitat factors. Region and grassland/forest were random factors. We selected the best model(s) supporting observed data – deltaAIC<4 in relation to other competing models – through Akaike Information Criteria (AIC – Burnham and Anderson, 2002). We tested residual normality and homoscedasticity prior to model selection, and evaluated/corrected for overdispersion and outlier influence only on the best model. When none model was the most plausible or null model was the best model, we performed model averaging over the full set of models. First, we analyzed data from all transects (both disturbed and undisturbed; n=144), and then reanalyzed data removing the sixteen transects from undisturbed sites (site 7, Fig. 1; n=128) since undisturbed and disturbed ecotones have different habitat structures (Appendix A1b, A1c). As variables were standardized, we interpreted variable slopes as standardized effect size (a significant effect when confidence intervals do not include zero). Analyses were conducted with glmmADMB, MuMIn, lme4, SYNCSA and vegan packages on R 3.1.3.
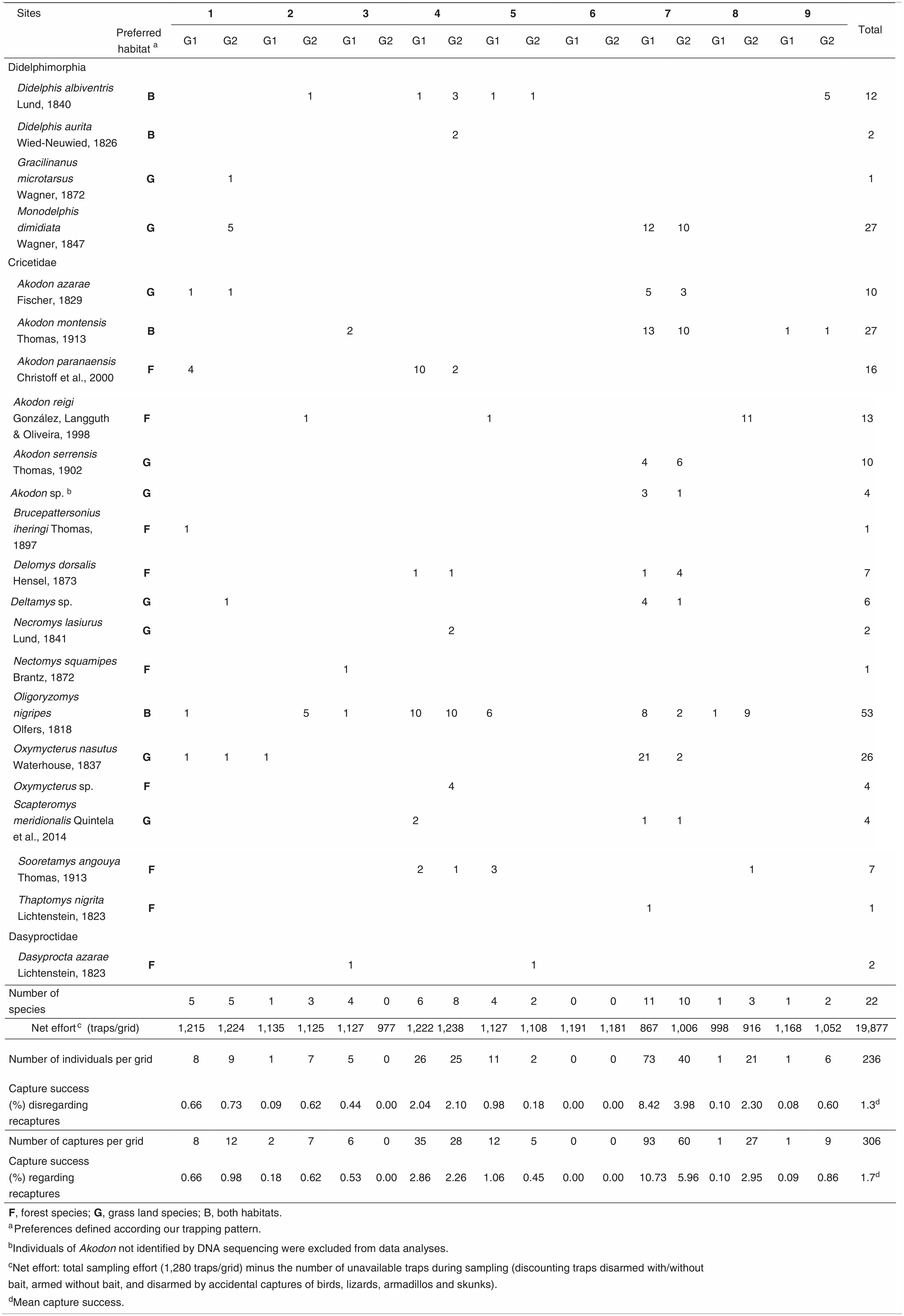
ResultsAn overall sampling effort of 19,877 trap-nights yields a mean success rate of 1.7% by site – 306 captures of 236 individuals, presenting higher success in the undisturbed site (Table 1). Seventeen species belonged to the order Rodentia and four to Didelphimorphia (Table 1). We sampled a total of 21±2.19 species (Jackknife 1=25.96±2.18; Chao 1=22.5±2.23) and recorded an average of 1.31±1.41 species and 1.97±3.1 individuals per transect. The most abundant species were Oligoryzomys nigripes Olfers, 1818, Monodelphis dimidiata Wagner, 1847, Akodon montensis Thomas, 1913 and Oxymycterus nasutus Waterhouse, 1837 (Table 1).



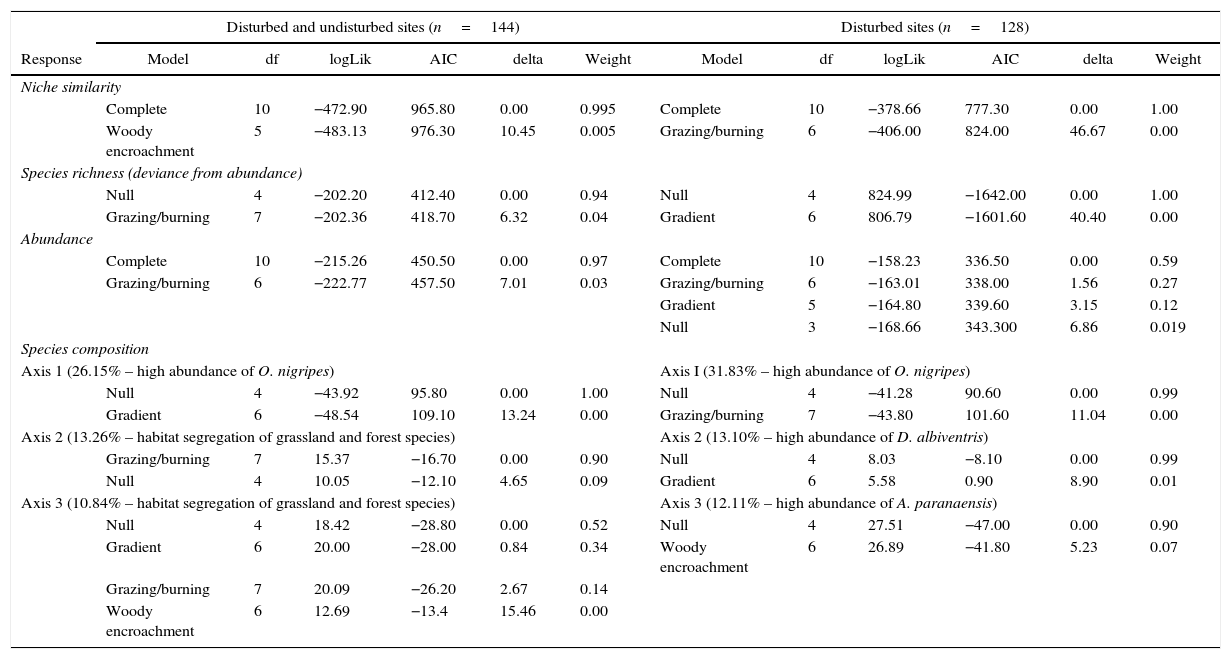
Mammal occurrence in all transects (both disturbed and undisturbed) supported the complete model as the most plausible for niche similarity and sample abundance (Table 2). Increases of around 215m in the distance from hydric resources decrease 0.78% niche similarity, and increases of 15% in the frequency of small forest patches on grasslands decreased 0.5% niche similarity (Fig. 2; Table S2 – Appendix A5). Assemblages from Brazilian Highland Grasslands had higher niche similarity in comparison to other regions (Table S2 – Appendix A5). Increases of 17.6cm in herbaceous vegetation height caused an increase of 1.5 individuals on average; canopy openness caused only a subtle decrease in abundance (Fig. 2). Assemblages from Brazilian Highland Grasslands were more abundant than from other regions (Table S5 – Appendix A5). The null model was supported by species richness and species composition on axes I and III (Table 2).
Relative importance of gradients of conditions and resources, grazing/burning, and woody encroachment in explain changes on the structure of small mammal assemblage across grassland–forest ecotones, Campos Sulinos – southern Brazil. Most plausible models, AIC difference between models (Delta) and Akaike weight are shown. Details of species composition analysis in Appendix A4. Complete results on Appendix A5.
| Disturbed and undisturbed sites (n=144) | Disturbed sites (n=128) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Response | Model | df | logLik | AIC | delta | Weight | Model | df | logLik | AIC | delta | Weight |
| Niche similarity | ||||||||||||
| Complete | 10 | −472.90 | 965.80 | 0.00 | 0.995 | Complete | 10 | −378.66 | 777.30 | 0.00 | 1.00 | |
| Woody encroachment | 5 | −483.13 | 976.30 | 10.45 | 0.005 | Grazing/burning | 6 | −406.00 | 824.00 | 46.67 | 0.00 | |
| Species richness (deviance from abundance) | ||||||||||||
| Null | 4 | −202.20 | 412.40 | 0.00 | 0.94 | Null | 4 | 824.99 | −1642.00 | 0.00 | 1.00 | |
| Grazing/burning | 7 | −202.36 | 418.70 | 6.32 | 0.04 | Gradient | 6 | 806.79 | −1601.60 | 40.40 | 0.00 | |
| Abundance | ||||||||||||
| Complete | 10 | −215.26 | 450.50 | 0.00 | 0.97 | Complete | 10 | −158.23 | 336.50 | 0.00 | 0.59 | |
| Grazing/burning | 6 | −222.77 | 457.50 | 7.01 | 0.03 | Grazing/burning | 6 | −163.01 | 338.00 | 1.56 | 0.27 | |
| Gradient | 5 | −164.80 | 339.60 | 3.15 | 0.12 | |||||||
| Null | 3 | −168.66 | 343.300 | 6.86 | 0.019 | |||||||
| Species composition | ||||||||||||
| Axis 1 (26.15% – high abundance of O. nigripes) | Axis I (31.83% – high abundance of O. nigripes) | |||||||||||
| Null | 4 | −43.92 | 95.80 | 0.00 | 1.00 | Null | 4 | −41.28 | 90.60 | 0.00 | 0.99 | |
| Gradient | 6 | −48.54 | 109.10 | 13.24 | 0.00 | Grazing/burning | 7 | −43.80 | 101.60 | 11.04 | 0.00 | |
| Axis 2 (13.26% – habitat segregation of grassland and forest species) | Axis 2 (13.10% – high abundance of D. albiventris) | |||||||||||
| Grazing/burning | 7 | 15.37 | −16.70 | 0.00 | 0.90 | Null | 4 | 8.03 | −8.10 | 0.00 | 0.99 | |
| Null | 4 | 10.05 | −12.10 | 4.65 | 0.09 | Gradient | 6 | 5.58 | 0.90 | 8.90 | 0.01 | |
| Axis 3 (10.84% – habitat segregation of grassland and forest species) | Axis 3 (12.11% – high abundance of A. paranaensis) | |||||||||||
| Null | 4 | 18.42 | −28.80 | 0.00 | 0.52 | Null | 4 | 27.51 | −47.00 | 0.00 | 0.90 | |
| Gradient | 6 | 20.00 | −28.00 | 0.84 | 0.34 | Woody encroachment | 6 | 26.89 | −41.80 | 5.23 | 0.07 | |
| Grazing/burning | 7 | 20.09 | −26.20 | 2.67 | 0.14 | |||||||
| Woody encroachment | 6 | 12.69 | −13.4 | 15.46 | 0.00 | |||||||
 measured in sites under different disturbance regimes. Inside each regression plot, we indicate the model from which variable comes. Dashed rectangles circumvent results disregarding undisturbed sites. Lines of predicted values and confidence intervals are shown. Niche similarity was transformed in percentage to analyze with Poisson GLMM.")
Relationship between structure of small mammal assemblages and environmental factors (deviations from the average) measured in sites under different disturbance regimes. Inside each regression plot, we indicate the model from which variable comes. Dashed rectangles circumvent results disregarding undisturbed sites. Lines of predicted values and confidence intervals are shown. Niche similarity was transformed in percentage to analyze with Poisson GLMM.
Grazing/burning effect was supported by species composition on Axis 2 (Table 2), which separate forest (− scores) from grassland species (+ scores) (biplots – Appendix A4). Increases in litter depth increased frequency and abundance of species that occur preferentially at grasslands, while increases in tree density promote the occurrence of mostly forest species (Fig. 2); random factors explained a low variance of species composition (Table S8 – Appendix A5).
Disregarding undisturbed ecotones (two grids [16 transects], two exclusive species, 113 individuals – Table 1), data suggest the complete model as the most plausible for niche similarity (Table 2). For niche similarity, distance from hydric resources, litter depth and small forest patches caused an effect 30–50% superior to other variables (Table S11 – Appendix A5). Distancing of 225.5m from hydric resources decreased 0.76% of niche similarity; increases of 1.1cm on litter depth decreased 0.5% of niche similarity. Quite similar to undisturbed sites, 15% increases in the frequency of small forest patches decreased 0.4% of niche similarity (Fig. 2). Abundance suggests both complete and grazing/burning models as equally plausible; however, no variables had significant effect (Table S13 – Appendix A5). The null model was supported by species richness and composition on disturbed sites (Table 2).
DiscussionConsiderable effort has been employed to understand the role of abiotic gradients, disturbances and woody encroachment for diversity and dynamics of ecotones (Bond and Parr, 2010; Müller et al., 2012; Veldman et al., 2015). Here, we tested models addressing these processes and perceived that mammal distribution and diversity depend on the spatial context, disturbance level and key microhabitat structures. We perceived habitat selection by forest and grassland species when combining data from both disturbed and undisturbed sites. When we removed undisturbed sites (i.e., more favorable conditions for specialist species), habitat selection was not evident because assemblages are species poor and functionally simplified in disturbed habitats with low carrying capacity.
Results showed that diversity and niche similarity in mammal assemblages depend on the spatial context where ecotones are inserted (e.g. region, availability of refuge habitats). We detected higher niche similarity and abundance in Brazilian Highland Grasslands than in other regions, and determined that mammal diversity on disturbed and undisturbed sites depends on microhabitat conditions provided by adjacent hydric resources. Hydric resources are acknowledged as refuges where species maintain stable and resilient populations (Milstead et al., 2007). Sites distant from water sources harbored species performing only a limited set of ecological functions, which highlight that only few non-volant small mammals are able to deal with adverse microclimatic conditions such as excessive dryness. Conversely, increases in the frequency of small patches and in litter depth means that these habitat structures can allow the co-occurrence of dissimilar species that may perform many ecological roles.
Evidence indicates that livestock management affects habitat selection by small mammals at Campos Sulinos (Pedo et al., 2010; Sponchiado et al., 2012); our study shows grazing/burning as relevant for species composition. Habitat selection by individual species alters assemblage structure as result of niche-related processes – differences in ecological requirements along gradients of habitat suitability and heterogeneity (Kingston and Morris, 2000; Wilson et al., 2010). Habitat selection by specialist species was evident only when combining undisturbed and disturbed ecotones. On highly disturbed sites (that allow occurrence of grass-like vegetation), opportunist species as Oligoryzomys nigripes and Didelphis albiventris occupy the entire gradient; the lack of grassland/forest specialists nullifies composition differences between grasslands and forests. Burning and intensive cattle grazing/trampling harm litter and vegetation structure – strata of plant senescent biomass, tussock herbs, shrubs, and trees (Matlack et al., 2001; Fox et al., 2003). Intensive management precludes the occurrence of grassland specialist species as Monodelphis dimidiata and Oxymycterus nasutus, which reach high densities on grassland habitats free or under light disturbances. These give security from predators and provide hidden places for foraging, digging shelters, resting, and nesting (Pedo et al., 2010; Sponchiado et al., 2012). Free-ranging cattle also alter the structure of forest understory and the soil cover. Ungulate presence in the forest interior depresses the litter layer and deteriorates understory structure through the foraging and trampling of seedlings, herbs and ferns (Dunstan and Fox, 1996); specialist forest species as Delomys dorsalis are not able to cope with sparse understory. Furthermore, increases in the litter layer provides shelter, nest sites and safety for movement of forest cursorial and semi-fossorial species (e.g. Thaptomys nigrita) (Clark and Kaufman, 1991). Therefore, increased disturbance intensity depresses the structure of mammal assemblages from both forest and grassland habitats, and the maintenance of simplified assemblages on disturbed sites may not guarantee functions performed by rich and functionally diverse assemblages.
In face of the current climate conditions, open ecosystems become subjected to forest advance mainly in regions with high precipitation rates (Behling et al., 2004). Data suggest that woody encroachment affected niche similarity, but this is not the main process shaping small mammal diversity in ecotones. Despite the weak relationship between species composition and woody encroachment, results suggest that woody encroachment impairs grassland species while benefiting forest ones (e.g. Akodon paranaensis). As forest patches are more similar to continuous forest – considering habitat and mammal species composition – than to the grasslands where patches are inserted, some forest mammals track changes in microhabitat conditions and explore habitats outside the forest edge. Our results also suggest that disturbances promote fauna replacement of grassland by forest species at higher rates than woody encroachment.
Our results highlight that specialist species are highly frequent on undisturbed ecotones, and that hydric resources are key structures that guarantee diversity in mammal assemblages. However, the refinement of our data may not be good enough to test the proposed hypothesis, and we acknowledge that an ideal study should sample a greater number of undisturbed ecotones (although such sites are each time more rare across landscapes). The actual scenario of high rates of habitat conversion and intensive livestock management makes mandatory conservation strategies that favor both managed and unmanaged areas distributed across landscapes (Andersen et al., 2012; Luza et al., 2014). Treated together, forest and grassy biomes form a highly diverse mosaic, and finding management thresholds that reconcile sustainable livestock production with conservation of grassland/forest fauna compose an exciting goal.
AuthorshipALL participated in data collection fieldwork, species identification (DNA sequencing), data analysis and manuscript preparation; GLG participated in data collection fieldwork, species identification (DNA sequencing) and manuscript preparation; VDP contributed in data analysis and manuscript preparation; SMH contributed in data analysis and manuscript preparation.
Conflicts of interestThe authors declare no conflicts of interest.
We would like to thank all landowners for granting access to their properties. We would like to acknowledge IBAMA (Brazilian Environmental Institute) (process # 30204-1), the Rio Grande do Sul State Environmental Bureau (process # 380) and the UFRGS Ethics Committee (process # 21539) for authorizing manipulation and collection of animals. We are grateful for the research grants received from CNPq and FAPERGS (# 11/2185-0). The first author received a master's scholarship from CAPES, VDP and SMH received research productivity grants from CNPq, and GLG received a postdoctoral fellowship from CNPq (156153/2011-4). We also thank Leandro D.S. Duarte, Marcus V. Cianciaruso, Jorge R. Marinho and Vinicius G. Bastazini for valuable suggestions during the course of the study. We thank the three anonymous reviewers for manuscript revision and Emma Bradley (International Master in Applied Ecology – IMAE) for English revision.
The following are the supplementary data to this article:

