





Climate change affects biodiversity in various ways but not linearly. Mountaintop species and ecosystems are expected to experience the strongest impacts of climate change. Lower tolerances to changes in habitat and decreased available surface area with increasing elevation should lead to smaller ranges and even extinctions of endemic species living on mountaintops. Thus, the potential impacts of climate change must be understood in order to achieve sound and long-lasting conservation and management of mountaintops. We evaluated the potential impacts of climate change on the distribution and conservation status of ten endemic bird species of eastern Brazil mountaintops based on ecological niche models. The models predicted a gradual reduction of suitable area for all species with a projected contraction of up to 94%, upwards shifts from 1039 to 1470m in elevation, and range shifts from 7 to 373km for 2070. Five groups of mountainous regions are highlighted as crucial for the current conservation of endemic birds. If these projections materialize, nine species of birds should be recognized as being under some level of threat. To diminish these effects, public policies and land conversion at local and regional scales and global reduction of greenhouse gas emissions are urgently needed.

There is strong evidence that climate change is impacting biodiversity, including the distribution and phenology of species (e.g. IPBES, 2019; Parmesan and Yohe, 2003; Vale et al., 2009). Although species generally present some niche plasticity, many are extremely fine-tuned to their habitats and considered too have limited abilities to cope with significant variation in habitat quality (Jiguet et al., 2007; Julliard et al., 2006). Augmenting pressures caused by global climate change are expected to increase the number of species potentially affected, causing various and unpredictable changes in community organization. While more specialized species should experience drastic reductions in their ranges, generalist species should spread into new available habitats (Clavel et al., 2010). The greatest impacts of climate change are predicted for species inhabiting high mountain ranges (e.g. Cuyckens et al., 2016; Sekercioglu et al., 2008), since montane species are more sensitive to climatic variation and the amount of appropriate surface area decreases with increasing elevation (Fernandes and Price, 1988; Rull and Vegas-Vilarrúbia, 2006).
Most predictions on climate change in mountainous ecosystems have been done for high mountain ranges, such as the Andes, the Alps and the Himalayas (e.g. Anderson et al., 2020; Carlson et al., 2017; Mavárez et al., 2018; Young et al., 2009). However, recent studies have shown that strong changes in habitat quality and area are also expected in smaller mountain ranges that are, in fact, far more numerous (e.g. Fernandes et al., 2016). Many smaller mountain ranges may represent ancient or geologically old formations that became eroded over time, as is the case of the Espinhaço, Mantiqueira and Serra do Mar mountain ranges in eastern Brazil, which are inhabited by some of the most diverse floras and faunas of the planet (for details see Fernandes, 2016; Fernandes et al., 2018; Silveira et al., 2016; Vasconcelos, 2011; Zappi et al., 2017).
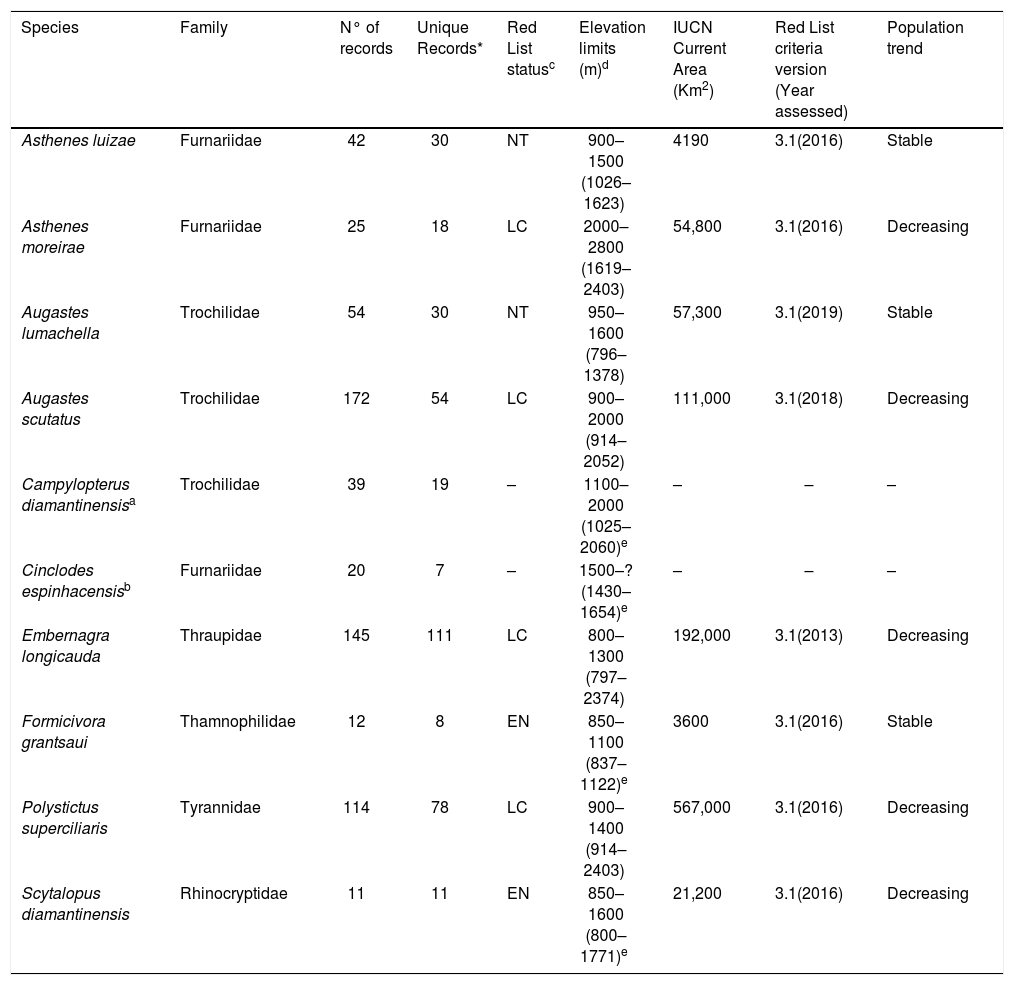
Despite the great diversity of birds in these three Brazilian mountainous regions (Vasconcelos and Rodrigues, 2010), the extent of potential impacts of climate change on species that inhabit these mountaintops has been poorly addressed (e.g. Marini et al., 2009a; Marini et al., 2010; Peterson et al., 2001; Vale et al., 2009). Ten bird species with different ranges are endemic to mountaintops in eastern Brazil (see Table 1) (Chaves et al., 2014; Vasconcelos and Rodrigues, 2010), of which two have yet to have their conservation status evaluated, four are listed as ‘Least Concern’, two as ‘Near Threatened’ and two as ‘Endangered’ due to their restricted distributions and severe habitat loss (IUCN, 2019). Additionally, since the methods used to estimate the distribution of these species (Minimum Convex Polygon – MCP) often incorporate large areas of unfavorable habitats (e.g. Ridgely and Tudor, 2009), their already reported distributions may be overestimates. This is because distributions determined by connecting occurrence points include valleys and wide lowland areas, which are actually not occupied by these species. Therefore, a detailed evaluation of the current and future distributions of these species is of major importance given the increasing threats driven by relentless climate change and land use conversion in these megadiverse mountains of eastern Brazil (Fernandes et al., 2018), and because sound conservation strategies must be designed now to better protect them into the future.
List of endemic bird species of eastern Brazilian mountaintops with the number of records, IUCN Red List status (2020), minimum elevation limits of occurrence, IUCN current Extent of Occurrence, IUCN Red List criteria version with year assessed and population trends.
| Species | Family | N° of records | Unique Records* | Red List statusc | Elevation limits (m)d | IUCN Current Area (Km2) | Red List criteria version (Year assessed) | Population trend |
|---|---|---|---|---|---|---|---|---|
| Asthenes luizae | Furnariidae | 42 | 30 | NT | 900–1500 (1026–1623) | 4190 | 3.1(2016) | Stable |
| Asthenes moreirae | Furnariidae | 25 | 18 | LC | 2000–2800 (1619–2403) | 54,800 | 3.1(2016) | Decreasing |
| Augastes lumachella | Trochilidae | 54 | 30 | NT | 950–1600 (796–1378) | 57,300 | 3.1(2019) | Stable |
| Augastes scutatus | Trochilidae | 172 | 54 | LC | 900–2000 (914–2052) | 111,000 | 3.1(2018) | Decreasing |
| Campylopterus diamantinensisa | Trochilidae | 39 | 19 | – | 1100–2000 (1025–2060)e | – | – | – |
| Cinclodes espinhacensisb | Furnariidae | 20 | 7 | – | 1500–? (1430–1654)e | – | – | – |
| Embernagra longicauda | Thraupidae | 145 | 111 | LC | 800–1300 (797–2374) | 192,000 | 3.1(2013) | Decreasing |
| Formicivora grantsaui | Thamnophilidae | 12 | 8 | EN | 850–1100 (837–1122)e | 3600 | 3.1(2016) | Stable |
| Polystictus superciliaris | Tyrannidae | 114 | 78 | LC | 900–1400 (914–2403) | 567,000 | 3.1(2016) | Decreasing |
| Scytalopus diamantinensis | Rhinocryptidae | 11 | 11 | EN | 850–1600 (800–1771)e | 21,200 | 3.1(2016) | Decreasing |
Previously treated as Cinclodes pabsti, but recently described as a new species (Freitas et al., 2012).
Ecological niche models have been extensively used to explore possible changes in species distributions caused by climate change (Araújo et al., 2011; Marini et al., 2009a, 2009b). They are also essential for understanding the persistence and distribution patterns of species over the long-term, thereby bolstering more efficient conservation strategies and efforts (Loyola et al., 2007). We propose to identify areas with high numbers of bird species endemic to mountaintops of eastern Brazil, evaluate the potential impacts of climate change on their distribution, present a list of the most threatened species and suggest urgent actions and policies for their conservation. We expect that the same trends observed for bird distributions in the high mountain ranges of the world will be mirrored in the lower mountains of eastern Brazilian (see Fernandes et al., 2016). Specifically, species distributions should shift to higher altitudes, and/or to higher latitudes, but with strong contraction in area due to the discontinuity of the eastern Brazilian mountainous areas to the south and the low altitude of the mountains (when compared to the Andes for example). Under these possibilities, we believe that the southern portion of the eastern Brazilian mountaintops, located at higher latitudes and subject to cooler climates, will be the most stable and relevant areas to maintain some of the endemic mountain birds.
Since many threatened species possess restricted distributions, we aim to evaluate the percentage of change in potential distribution area for future scenarios in relation to the current projection. The results will allow us to identify species that may need special conservation attention if these contraction and expansion percentages are applied to the distribution areas used by IUCN to determine the conservation status of species. Considering that the species of interest already have restricted distributions, we expect that all ten of them will experience contractions in their distribution areas in relation to the literature, and as such should have their conservation statuses reviewed anew.
MethodsStudy areaThe study area is located in eastern Brazil (37°–49°W, 9°–25°S), and consists of the areas above 800m of three mountain ranges (Espinhaço, Mantiqueira, and Serra do Mar) defined by a 300km buffer area around all the occurrence records of all of the ten studied species (Supplementary Fig. S1). Therefore, the global study area encompasses the individual model-projection areas for each species, which is restricted to a buffer area around their occurrence records (Supplementary Fig. S2) (for details see Supplementary Text S1).
Species ecological niche modelsEcological niche models (ENMs) are based on known presences and absences or pseudo-absences (PAs) (Peterson et al., 2011). Thus, occurrence records for the ten bird species endemic to mountaintops of eastern Brazil were compiled from the literature, field observations and museum specimens whenever geographic coordinates were available (see Vasconcelos, 2008) (Table 1, Supplementary Fig. S2, Table S1). Bird species identification followed the systematics and nomenclature of the latest revision of the Brazilian Ornithological Committee (Piacentini et al., 2015). True absence records were replaced by randomly generated PAs. For details of key steps for model building and analysis see Supplementary Box S1 with ODMAP (Overview, Data, Model, Assessment and Prediction) protocol (according to Zurell et al., 2020), and Supplementary Text S1.
Because many climatic variables, after elimination of those that are highly correlated (negatively or positively), can present a certain level of collinearity, and so would negatively affect ENMs (e.g. De Marco and Nóbrega, 2018), we selected two sets of predictor variables for building ENMs. The first set was formed by the least correlated climatic variables (r Pearson<0.7) and the second by Principal Component Analysis (PCA) axes (for details see Supplementary Text S1, Table S2). These sets of predictor variables were obtained for the current and future scenarios with approximately 1km2 resolution for each species. For future scenarios we considered two periods (until 2050 and until 2070) with four Representative Concentration Pathways (RCP: rcp2.6, rcp4.5, rcp6.0, rcp8.5) (see IPCC, 2013) and three Global Climate Models (GCMs: CCSM4, HadGEM2-ES, MIROC-ESM), resulting in a set of 24 scenarios per bird species.
Data were partitioned into ‘calibration’ and ‘test’ sets using the ENMeval package (Muscarella et al., 2014). For species with>25 occurrences, the data were partitioned ten times, four sets with ‘block’ methods and five with ‘randomkfold’, each set with 75% of data for calibration and 25% to test. The last set consisted of all records for calibration and testing. For species with <25 occurrences, we use the ‘jackknife’ (n-1 jackknife) method, by which each of n occurrence localities is used for testing, while all others are used for calibration (for details see Supplementary Text S1). For each data partition set and each predictor set, ENMs were created using five algorithms (GAM, GBM, GLM, MAXENT, and RF) implemented with the Biomod2 framework (Thuiller et al., 2009) in the R programming environment (RStudio Team, 2019). Model performance for species with >25 records was assessed based on the average True Skill Statistic (TSS) (Allouche et al., 2006) where models with TSS>0.80 were selected (see the Biomod2 manual for model selection for consensus building). For model evaluation for species with <25 records we computed the p-value (p<0.05) as proposed by Pearson et al. (2007).
For each bird species, the selected models for each predictor set were projected for current and future scenarios, considering a restricted region (see Supplementary Fig. S2). The projections were transformed into binary maps using the maximum sensitivity and specificity threshold for species with more than 25 records. For species with less than 25 records, the lowest presence value was used as a limit for occurrence. The same estimated and used threshold for the current scenario was used for each projection in future scenarios.
AnalysisFor the two sets of predictor variables, we estimated the current and future distributions of each species through the sum of the binary maps resulting from the selected models. We considered the species present in a cell with 50 percent or more projection frequency (for details see Supplementary Text S1). To identify areas of uncertainty (model extrapolation) in projections over future scenarios (see Owens et al., 2013), we performed a Mobility-Oriented Parity (MOP) metric using the kuenm package in R (Cobos et al., 2019). To determine if there was a difference between the areas projected by the models of the two sets of predictor variables, we performed the Wilcoxon paired test for non-parametric data.
To determine the areas with the highest predicted richness of endemism for each scenario, we combined the binary maps with the projected area of each species. To evaluate changes in species richness, the richness maps of each future scenario were superimposed on the current scenario to assess changes in environmental suitability for at least one species, indicating stable areas with expansion and retraction (see Supplementary Text S1). For each scenario and for each richness class, we estimated the area occupied, the average altitude and the changes between the current and future scenarios (see Supplementary Text S1).
To assess potential impacts of climate change individually for each species, we overlapped the current projection of each against each of its future scenarios and estimated the percentage change in area size, distance and direction of displacement from the center of the distribution area, and the variation in mean altitude between the current scenario and each future scenario. To evaluate the possibility of reevaluating the conservation status of the studied species, the percentage change in the range of each species between the scenarios was applied over the distribution area estimated by the IUCN. All analyses were performed in the R environment (RStudio Team, 2019) (see Supplementary Text S1).
ResultsA total of 634 records were compiled for the ten species (Table 1), and the total number of models created for each set of predictor variables was 565, ranging from 35 to 90 per species (Supplementary Table S3). The number of selected models (TSS>0.80) for projections and final consensus for each species ranged between 5 and 70 for climatic variables as predictors and from 10 to 72 for PCA axes as predictors (Supplementary Table S3). The areas of uncertainty or extrapolation predicted by the MOP were larger for climate data as predictors than for PCA axes as predictors. For the two sets of predictor variables, the areas denominated with uncertainties practically did not overlap with predicted suitable areas (Fig. 1). Since the projected area did not overlap with areas of uncertainty, we compared the extent of this area produced by the models of the two sets of predictor variables and did not observe any significant difference (p=0.2394).
, in relation to the values of 2050 and 2070 and different scenarios of gas emissions (RCP), considering two sets of predictor variables (red color indicates loss of suitable conditions for all species; green color indicate stability and blue gain of range for one or more species). Lines two and four show the results of the Mobility-Oriented Parity (MOP) analysis, which indicates areas of uncertainty for one or more species (with possibility of model extrapolation) (see Cobos et al., 2019; Owens et al., 2013).")
Changes in richness and distribution of bird species endemic to eastern Brazilian mountaintops, current scenario (first column, lines 1 and 3), in relation to the values of 2050 and 2070 and different scenarios of gas emissions (RCP), considering two sets of predictor variables (red color indicates loss of suitable conditions for all species; green color indicate stability and blue gain of range for one or more species). Lines two and four show the results of the Mobility-Oriented Parity (MOP) analysis, which indicates areas of uncertainty for one or more species (with possibility of model extrapolation) (see Cobos et al., 2019; Owens et al., 2013).
For both groups of predictor variables, the richness maps produced by the projection of the models indicated a gradual and marked reduction between scenarios in suitable area for mountaintop endemic birds, as well as an altitudinal shift of species richness. Current projections indicate that mountaintop endemic birds are distributed over an area of 149,930 and 123,152 km2 and an average altitude of 1039.1 and 1046.2m, considering projections with climate data and PCA axes, respectively, as predictor variables (Supplementary Table S4). The greatest species richness (6 species) was predicted for an area of 150 km2 restricted to the highest altitudes (>1519m) (Fig. 2). The projections of future scenarios suggest a decrease in the number of species in the richest centers (a maximum of 5 species) and a contraction in the suitable area for richness maintenance, ranging between 72 and 85% for the RCP scenario for 2050 and between 80 and 94% for 2070, considering the two predictor sets (Supplementary Table S4). Similarly, an altitudinal displacement is predicted with mean altitudes of 1278 and 1289m by 2050 and 1458 and 1470m by 2070, considering the two sets of predictor variables. (Supplementary Table S4). New suitable area for richness expansion of endemic bird species ranged between 10 and 3020km2 (Supplementary Table S4), for the different future RCP scenarios.
, with different scenarios of gas emissions (RCPs), considering Ecological Niche Models (ENMs) built with two predictor sets (climatic variables and PCA axes).")
Changes in area occupied and average altitude by different richness classes of endemic birds of eastern Brazilian mountaintops, for the scenarios of 2050 and 2070 (lines of graphs), with different scenarios of gas emissions (RCPs), considering Ecological Niche Models (ENMs) built with two predictor sets (climatic variables and PCA axes).
The reduction and displacement of the distribution of species result in changes in the spatial pattern of species richness, which is unevenly distributed throughout the study area. The current projections suggest the existence of five groups of mountainous areas with conditions for the presence of endemic mountaintop bird species: (a) north-central Bahia; (b) isolated areas in northern Minas Gerais and southern Bahia; (c) northern Serra da Mantiqueira; (d) south-central portion of the Espinhaço Range; and (e) small, isolated areas in western Espírito Santo (Fig. 3).
, based on models with climatic variables as predictors; hatched areas indicate five groups of highlands relevant for endemism: (a) north-central part of the state of Bahia with a species richness hotspot in the Chapada Diamantina; (b) mountains in northern Minas Gerais and southern Bahia; (c) northern Serra da Mantiqueira; (d) the largest block extending from the central-south portion of the Espinhaço Range; and (e) a group of small, isolated areas in western Espírito Santo, particularly in Serra do Caparaó.")
Predicted species richness of endemic birds of eastern Brazilian mountaintops (highest species richness in red, single species in blue), based on models with climatic variables as predictors; hatched areas indicate five groups of highlands relevant for endemism: (a) north-central part of the state of Bahia with a species richness hotspot in the Chapada Diamantina; (b) mountains in northern Minas Gerais and southern Bahia; (c) northern Serra da Mantiqueira; (d) the largest block extending from the central-south portion of the Espinhaço Range; and (e) a group of small, isolated areas in western Espírito Santo, particularly in Serra do Caparaó.
Five major groups of mountains (Fig. 3) will lose species (red areas, Fig. 1), with the loss of practically all conditions to support the endemic species in Chapada Diamantina and northern Minas Gerais. The other three areas in the south-central portion of the Espinhaço Range would experience a decrease in area, but also the presence of a center with adequate stability, especially in the Iron Quadrangle (green area in Fig. 1).
Considering the species individually, for the two sets of predictor variables, we observed a trend for a gradual reduction in the projected distribution for future periods and each RCP scenario for all the species (Supplementary Table S5, Fig. S3). The gain of new areas with suitable conditions for each species in future periods and different RCP scenarios varied, for the model predictor sets (climatic variables and PCA axes) between a maximum of 3400 and 2500km2 for 2050 and 900km2 and 2500 for 2070 (Supplementary Table S5). The projected distribution area for each species, considering the two sets of predictor variables (climatic variables and PCA axes), exhibited a maximum elevation shift of 664 and 607m for the RCP scenario of 2050 and 694 and 744m for 2070 (Fig. 4), respectively. We found a tendency for a southwest/west-southwest latitudinal shift of the center of the distribution area, with a maximum shift under RCP scenarios of 324 and 318km for 2050 and 373 and 342km for 2070, for climatic data and PCA axes as predictors, respectively (Fig. 4, Supplementary Fig. S4 and Table S6).
 and latitudinal (km) range shifts for each species considering models built with two sets of predictors (climatic variables and PCA axes) and projected in four RCP scenarios of for the scenarios of 2050 and 2070. Light red represents the 2050 scenario and light green represents the 2070 scenario. Values for the RCP bars, of both altitudinal and latitudinal shift panels, indicate the average difference for the IUCN data and the projections of the current scenario. Zeros for RCP bars indicate that the species did not have a projected area with suitability for that scenario and did not present altitudinal and/or latitudinal range shifts.")
Projected altitudinal (m) and latitudinal (km) range shifts for each species considering models built with two sets of predictors (climatic variables and PCA axes) and projected in four RCP scenarios of for the scenarios of 2050 and 2070. Light red represents the 2050 scenario and light green represents the 2070 scenario. Values for the RCP bars, of both altitudinal and latitudinal shift panels, indicate the average difference for the IUCN data and the projections of the current scenario. Zeros for RCP bars indicate that the species did not have a projected area with suitability for that scenario and did not present altitudinal and/or latitudinal range shifts.
The percentage change in the projected area for the current scenario of each species relative to future scenarios indicated a gradual reduction ranging from 44.5% to 100% from the milder RCP scenario (2.6) to the most drastic (8.5) for both 2050 and 2070, but more pronounced in 2070 (Supplementary Table S5). When the percentage of predicted change for the different scenarios was projected over the distribution area used to determine the current conservation status of the species by the IUCN (2018), all species were indicated as in need of conservation status reevaluation at some point, with the exception of P. superciliaris (Supplementary Table S7). The species C. diamantinensis and C. espinhacensis could not be evaluated since both are considered subspecies by the IUCN. The species Formicivora grantsaui and S. diamantinensis will lose their total distribution area under most of the predicted scenarios.
DiscussionAs we expected for the climate change scenarios examined, we found a pattern of reduced area and altitudinal displacement for endemic birds of mountaintops in eastern Brazilian, which is similar to that projected for bird species of higher mountain ranges in temperate and tropical regions, such as the Andes (e.g. Freeman et al., 2018; Popy et al., 2010). However, consequences may be even more severe here, as the mountains are low and spatially restricted, with no available areas for altitudinal or latitudinal displacement. Further, as demonstrated by Pena et al. (2017), parts of these areas are highly impacted by mining actions and vegetation changes caused by human-induced fires and invasion of exotic grasses (Alves and Silva, 2011; Aximoff et al., 2016; Carmo and Kamino, 2015; Conceição et al., 2015). Thus, with our projections the future prospects are not optimistic, suggesting dire perspectives for the species considered in the present study. In order to minimize this pessimistic scenario, our main recommendation is to invest in the creation of new protected areas, especially in the south-central portion of the mountains in eastern Brazil, since this region showed higher stability between the scenarios. On the other hand, our findings reinforce the strategic importance of smaller mountain ranges for preserving biodiversity and provisioning ecosystem services (Callisto et al., 2019; Silveira et al., 2019). The upward shift of the species’ elevational range, due to climate change will mainly affect mountain endemic species, whose greatest richness will be concentrated in the highest areas, according to our projections.
The MOP analysis (Cobos et al., 2019; Owens et al., 2013) indicates large areas of uncertainty and extrapolation for the projection of models under future scenarios using climate variables as predictors in comparison to using the PCA axes. However, the identified areas of uncertainty practically do not overlap with areas projected as adequate by the two data sets, so as not to influence the results. Thus, the results from using the two sets of predictor variables to build the ENMs can be compared and produced projections that are not significantly different, making them complementary and reinforcing the results. The projections indicate that variation among the RCP scenarios can lead to different conclusions regarding trends in the distribution of richness and species for each period (2050 and 2070).
All species, throughout the periods and among different scenarios, experienced a displacement in their distributions to higher areas, as expected. As noted for variation in richness, these displacements also indicate a contraction in area due to the lack of available mountain area at higher elevation. This finding indicates the “escalator to extinction” effect, as previously proposed (see Freeman et al., 2018). In a meta-analysis, Parmesan and Yohe (2003) evaluated the potential responses of organisms to climate change, among which a latitudinal displacement in the face of temperature increase would be expected. In the present study, a latitudinal displacement to the southwest/west-southwest (i.e., to higher latitudes) is supported by the existence of potential areas where temperatures are milder. On the other hand, a gradual increase in latitudinal displacement for the species studied here between climate change scenarios (from the mildest to the most drastic RCPs) was only partially observed (see Fig. 4). The limited latitudinal displacement, without a gradual increase, as observed for all species that did not have their areas go extinct, may be due to two main reasons. First, to make estimates closer to reality we determined a movement area consisting of a buffer area with a radius ranging from 30 to 300km around the occurrence records for each species (see Brown and Yoder, 2015; Thuiller et al., 2009). For some scenarios, the range shift of some species is less than the radius of the buffer area (see Table S6). Hence, this delimitation reduces over-projections in areas where the species does no currently occur, which is important when we assess changes in occupied area to identify possible conservation needs. Therefore, this delimitation, at least partially, restricts the latitudinal shift. Second, the limited response recorded could also have been influenced by the fact that these mountains do not extend continuously across a broad latitudinal gradient, as observed in the Andes for instance. This indicates that temperature variation along the altitudinal and latitudinal gradient has a strong influence on the prediction of species distributions and on the determination of bird species conservation statuses. In addition, the conservation status of each species may be influenced by their specific dispersal ability, habitat suitability and ecological interactions, which remain unknown for most of the studied species. Therefore, special attention should be directed to conducting detailed studies on the ecology and natural history of these bird species.
The few studies in Brazil that have modeled the distributions of bird species under climate change scenarios reached similar conclusions. Marini et al. (2010) projected a 47% contraction in the area for Amazona pretrei, while Souza et al. (2011) projected a 45% reduction in average area of 44 species of endemic birds of the Atlantic Forest. To evaluate the effectiveness of conservation units for the protection of birds of the Cerrado biome under scenarios of climate change, Marini et al. (2009a) modeled the distribution of 38 species and observed a contraction in their areas, concluding that the conservation units are not sufficient for their conservation. Although Marini et al. (2009a) included the species A. scutatus, A. lumachella, E. longicauda, A. luizae and P. superciliaris, the estimated distribution area of these species in our study is greater, perhaps due to their use of variables at a different resolution. Marini et al. (2009b) projected changes in distribution area for 26 bird species of the Cerrado biome, varying from an increase of 5% to a contraction of up to 80%, plus a trend of displacement to the southeast in future scenarios of climate change, regardless of species dispersal ability. This variation was less apparent in our study because the species responded idiosyncratically to possible changes, which was heavily influenced by the availability of high-elevation areas, which are correlated with, and influence, the predictors of the distribution models.
Among the areas with higher concentrations of endemic species, and which are likely to remain stable under future scenarios, we emphasize the importance of the following regions for both conservation and future research: the Chapada Diamantina region and the south-central Espinhaço Range (including the Iron Quadrangle). The area from the south-central portion of the Espinhaço Range to the Serra da Mantiqueira and Serra do Mar (green areas in Fig. 1; Fig. 3c and d) can be considered to contain the most important mountainous areas for conservation and research of the endemic mountaintop birds of eastern Brazil because this region is expected to maintain, at least in part, a higher concentration of endemic species, and is likely to remain stable for some of the species according to the future scenarios. Furthermore, small but new areas with environmental suitability under future scenarios and, as they are within the current range of species’ distributions, they have the potential to be colonized. However, both rupestrian and high-altitude grasslands in the region are under great anthropic pressure, such as frequent fires, mining, conversion into pastureland, urban sprawl, road construction and biological invasion by exotic plants (see Fernandes et al., 2014; Pena et al., 2017; Stattersfield et al., 1998). Therefore, with these conditions, the region cannot fulfill its role as a refuge for endemic bird species under scenarios of climate change, with the exception of small, adjacent areas that may be less affected, such as Santuário do Caraça Private Reserve, Serra do Gandarela National Park and Serra do Rola Moça State Park, despite this last unit being peri-urban and subjected to intense human disturbances.
Even if the factors responsible for the loss of natural habitats are managed in the future, a reduction of between 72 to 94% in the current area suitable for the occurrence of at least one endemic species is expected due to climate change (Supplementary Table S4). Therefore, climate change can be an important factor in reducing the size of the area of occurrence of bird species endemic to mountaintops in eastern Brazil, reinforcing the need for research and the application of conservation and management strategies in the present time. To diminish these effects, public policies and land conversion at local and regional scales and global reduction of greenhouse gas emissions are urgently needed.
If we project the percentages of change observed between the current scenario and future scenarios on the Extent of Occurrence (EOO) (Supplementary Table S7) adopted by the IUCN to determine the current conservation status of species, most species would need to have their conservation status reevaluated and many species would have to be included in some category of threat under Criterion B of the IUCN. The impacts of the projected climate change on EOO indicate that seven to eight species of the ten studied species may require a reevaluation by 2050, according to the RCP scenario, and eight to nine species by 2070. The only species that would be out of any level of threat would be P. superciliaris, not because projection results indicate that there would be no changes, as they were relatively high, reaching up to a 93% reduction (Supplementary Table S7), but because the EOO of the IUCN for the species is considerably extensive, encompassing wide areas where the species is not currently known (based on the authors’ field observations). Furthermore, by 2070 the EOO remaining projected for A. lumachella is <100km2 of suitable environment, and thus the species could be re-evaluated as Critically Endangered. Some species, such as F. grantsaui and S. diamantinensis, may come to an end, according to the vast majority of scenarios, due to the loss of environmental suitability throughout their known ranges. Furthermore, species such as C. espinhacensis and C. diamantinensis, which are not officially recognized as full species by some committees, such as the IUCN, have projected changes of over 99% for all scenarios, indicating that they may lose their entire area of distribution before they are finally recognized by some taxonomy committees. This is a worrying finding that stresses the importance of taxonomic research in support of solid conservational actions.
The species that need greater attention in relation to studies on their biology and population monitoring are F. grantsaui, A. lumachella, S. diamantinensis, C. espinhacensis and C. diamantinensis, because their distributions are restricted to the Chapada Diamantina and center-north Espinhaço, areas that may be adversely affected by climate change and land use conversion. On the other hand, given their projected reduced distributions, and the central point of their distributions being located in the south-central portion of the Espinhaço and Mantiqueira ranges, populations of A. moreirae and A. luizae should be monitored along elevation gradients. These observations, based exclusively on the change projected for EOO, indicate the need for population and habitat use studies to produce solid information on species conservation status.
In summary, the models developed here clearly show that these endemic birds will have their geography drastically altered, including reductions in suitable area. These species will need to migrate to higher altitudes in the mountain landscape, be displaced southward, and forced to move away from their actual distribution centers. Similar trends have been reported in other regions (Freeman and Freeman, 2014; Marris, 2007; Sekercioglu et al., 2008). Freeman et al. (2018) argued that “climate change-driven mountaintop extirpations may be more likely in the tropics, where temperature seems to exert a stronger control on species’ range limits than in the temperate zone” (see also Laurance et al., 2011). Our results support the expectations that species will face a greater contraction in their distribution because of decreased available area for altitudinal displacement due to the intermediate altitude of the mountains.
Our pessimism regarding the fate of endemic mountaintop birds is augmented, since potential displacements were not fully evidenced, because species latitudinal movements were practically stabilized between the periods and among RCPs, most probably due to the restricted distribution of the bird species studied and lack of area for latitudinal displacement. An additional factor that negatively influences this latitudinal displacement is the latitudinal discontinuity of elevated areas in the eastern Brazilian mountains. This geography differs from that of the Andes due to the existence of geographical barriers such as glaciers, large lakes and arid valleys, although the mountains extend from 0° to 50°S. To attempt to avoid the loss of bird species endemic to mountaintops in eastern Brazil, urgent and bold research and public policies in conservation, and the accomplishment of the commitments of sustainable development goals, with land use planning and urgent reduction of greenhouse gas emissions, are needed. Although this seems a far-off distant goal given the current reduction in Brazil's investment in science (Fernandes et al., 2017), and misguided policies in conservation (Pacheco et al., 2018), actions ought to be taken now if the focus is on biodiversity and a more sustainable world.
Conflicts of interestWe have no conflicts of interests to declare.
We thank I. Arias, J.P. Metzger, and three anonymous reviewers for their comments on earlier versions of this manuscript. This work was supported by the Brazilian National Research Council (CNPq) [grant number 500995/2011-7 PDJ] assigned to DH and supervised by GWF; and the Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG) assigned to GWF.
The following are supplementary data to this article:

