


Because of changes in riparian zones and water properties, human disturbances in terrestrial ecosystems can affect the decomposition of organic matter and invertebrate assemblages in forest streams. The aim of this study was to evaluate how changes in the riparian zones influenced leaf breakdown rates and colonization by invertebrate shredders. Leaves of Miconia chartacea were incubated in four streams of the same watershed that presented different conservation levels in the riparian zones. Leaf breakdown rates were higher in the presence of shredders and in the most preserved stream. In terms of abundance and biomass, shredders differed among streams, and the importance of these organisms on leaf decomposition decreased in altered streams. These results suggest the conservation level in the riparian zones influenced leaf decomposition mediated by shredders, and the observed decrease in breakdown rates was probably due to the high sensitivity of shredders to changes in the availability of food resources and habitat.
The increase in human population and environmental impacts has caused many changes in both terrestrial and aquatic ecosystems (Allan, 2004). Because of the linkages between these environments, changes in the riparian zones, such as deforestation to create grazing areas and the replacement of native species by monocultures (Vorosmarty et al., 2010), also affect aquatic communities and important ecological processes occurring in stream ecosystems (Landeiro et al., 2010).
Changes in the riparian zones can modify the quality and quantity of leaf litter produced (Silva-Junior et al., 2014). Given that allochthonous material is an important energy source to heterotrophic organisms (Vannote et al., 1980), changes in the inputs of leaf litter can influence aquatic invertebrate assemblages by modifying the distribution and abundance of different taxa (Masese et al., 2014) and functional feeding groups (Encalada et al., 2010). Among these, invertebrate shredders, which feed on leaves imported from the riparian vegetation, are mainly affected. Through their activity, these organisms transform the coarse particulate organic matter into fine particles, feces and dissolved organic matter, exerting an important role in the detritus chain (Cummins, 1973). Therefore, changes affecting the distribution of shredders can modify the availability of resources along the food chain and stream ecosystem functioning (Dudgeon, 2006).
Traditionally, the assessment of stream integrity used structural indicators, i.e., monitoring aquatic communities and the physical and chemical parameters of water (Callisto et al., 2004). More recently, the evaluation of ecological processes, such as leaf decomposition, has been used as an additional tool for stream assessment (Graça et al., 2015). The decomposition process is an important functional indicator of stream ecosystems (Gessner and Chauvet, 2002), as it comprises both the riparian vegetation and biotic and abiotic components of the aquatic environment (Cummins, 1974). The use of functional indicators is widespread in Europe and North America. In Brazil, few studies have used this approach (see Silva-Junior and Moulton, 2011; Silva-Junior et al., 2014), and the leaf decomposition process was normally used to compare the functioning of ecosystems that were clearly different, such as preserved and impacted streams (Gonçalves et al., 2014).
Herein, the leaf decomposition process was evaluated in four streams of the same watershed that presented different conservation levels in the riparian zones. The aim of this study was to evaluate how changes in the riparian zones influenced leaf breakdown rates and colonization by invertebrate shredders. Assuming that changes in the abundance and species composition of the riparian vegetation decrease the diversity and availability of leaf litter on streambed, we hypothesized that breakdown rates, and the importance of invertebrate shredders on this process, would decrease in altered streams, i.e., the most preserved stream would present the highest breakdown rates and invertebrate assemblages with high proportions of shredders in terms of abundance and biomass.
Materials and methodsStudy areaThe four streams studied (Norte, Luxemburgo, Macuco and Banana) belong to the Santa Maria da Vitória River Basin and are located in the State of Espírito Santo, Southeastern Brazil. Norte Stream (20°01′43.9″S–40°32′00.8″W) presents a high level of human disturbance. The native vegetation was completely removed in one of the margins, and a narrow line of trees remained along the other margin. Both margins had signs of erosion, and the predominant substrate types were sand banks and leaf patches. Luxemburgo (20°08′15.6″S–40°35′44.4″W) is the second most disturbed stream. Despite the presence of the riparian vegetation in both margins, exotic fruit trees (banana and jackfruit) replaced the majority of native species. The studied reach had a rocky substrate dominated by large sand banks and some leaf patches. Macuco (20°01′23.1″S–40°32′58.6″W) and Banana (20°02′22.1″S–40°31′53.9″W) streams are located in fragments of Atlantic Forest and are more preserved. However, a monoculture of Eucalyptus globulus Labill. (1000m2) and a small rural property were located close to one of the margins of Macuco. The streambed of these streams was heterogeneous, with the presence of pebbles, gravel and leaf patches. More information about the studied streams and the physical and chemical properties of water (Table S1) is available in the supplementary material.
Experimental designSenescent leaves of Miconia chartacea Triana (Melastomataceae) were collected in the riparian zone of Macuco Stream using 4 litter traps fixed at 1.5m height and dried at room temperature. This species was chosen because it was abundant in the riparian vegetation of the studied streams, and shredders were frequently observed consuming these leaves. Portions of 2.00±0.05g of leaves were placed in litter bags (10cm×15cm) of fine (0.5mm) and coarse (10mm) meshes and incubated in the four streams, in reaches with similar conditions of depth and flow. Thirty-six litter bags of each mesh were incubated per stream, totaling 288 samples. Four replicates of each mesh were collected in each stream at the intervals of 3, 7, 15, 30, 45, 60, 90, 120 and 150 days of incubation. Additional litter bags (time 0) were used to correct mass loses that occurred during sample handling and transport and to determine an oven dry mass/air dry mass correction factor.
In the laboratory, the leaves of each sample were washed over a 250μm mesh and dried (60°C, 72h) to determine the remaining mass. Invertebrates found in the coarse mesh bags were fixed in 70% ethanol and posteriorly identified and assigned to functional feeding groups using specific identification keys. Individuals were then separated by taxon, counted, dried (60°C, 72h) and weighed in an analytical balance (0.1mg) for the determination of biomass.
Data analysisA negative exponential model was used to evaluate the decomposition process. The regression of the natural logarithm of the proportion of the remaining leaf dry mass and the time elapsed (days) was calculated, and the regression slope was used as the breakdown rate (k). Separate models were used to calculate the breakdown rates in fine (kf) and coarse mesh bags (kc) and to compare them, as well as the kc/kf ratios, among the studied streams (Gessner and Chauvet, 2002). The differences in the taxa composition of shredders among the streams were tested by an Analysis of Similarities (ANOSIM), based on the abundance data. General linear models (GLMs) were used to compare the proportion of shredders in terms of abundance (number of individuals per litter bag) and biomass (total dry mass per litter bag) among streams and incubation times (ln[x+1] transformed data). We tested normality on residuals from GLMs for each response variable. When a model was significant, Tukey tests were used for post hoc comparisons among streams and incubation times. All of the statistical analyzes were performed using SPSS (version 23.0 for Macintosh, SPSS, Chicago, Illinois) and Primer 6 (Primer-E Ltd.).
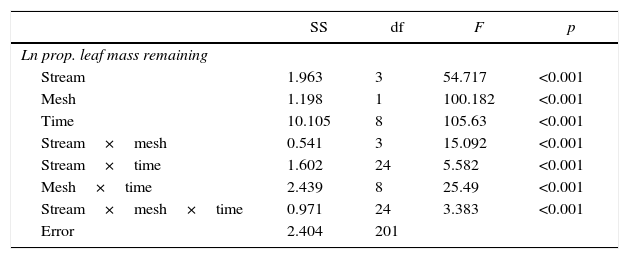
ResultsLeaf breakdown rates differed among streams, mesh sizes and incubation times (Table 1). The natural logarithm of the proportion of the remaining leaf dry mass decreased with time, and the lower mass losses were observed in fine mesh bags in all of the streams and also in coarse mesh bags in Norte Stream (Fig. 1). The coarse mesh bags incubated in Banana had the highest mass loses, while those incubated in Luxemburgo and Macuco had intermediate values. With the exception of Norte, the breakdown rates in coarse mesh bags (kc) were higher than the rates observed in fine mesh bags (kf) in each studied stream (Fig. 2). The importance of invertebrate shredders, as indicated by the kc/kf ratio, was higher in Banana (2.36) and decreased in altered streams (Macuco: 2.17, Luxemburgo: 1.75 and Norte: 0.96).
Variation in the natural logarithm of the proportion of leaf mass remaining depending on the stream (Norte, Luxemburgo, Macuco and Banana), mesh size (coarse or fine) and experimental time (in days) tested by general linear models. SS=sum of squares. df=degrees of freedom.
| SS | df | F | p | |
|---|---|---|---|---|
| Ln prop. leaf mass remaining | ||||
| Stream | 1.963 | 3 | 54.717 | <0.001 |
| Mesh | 1.198 | 1 | 100.182 | <0.001 |
| Time | 10.105 | 8 | 105.63 | <0.001 |
| Stream×mesh | 0.541 | 3 | 15.092 | <0.001 |
| Stream×time | 1.602 | 24 | 5.582 | <0.001 |
| Mesh×time | 2.439 | 8 | 25.49 | <0.001 |
| Stream×mesh×time | 0.971 | 24 | 3.383 | <0.001 |
| Error | 2.404 | 201 | ||
 and coarse mesh bags (dashed lines, open circles) on the elapsed time (days) at the Norte (A), Luxemburgo (B), Macuco (C), and Banana (D) streams.")
 of leaves of M. chartacea in coarse (empty bars) and fine mesh bags (solid bars) in the four studied streams. Breakdown rates with the same letter do not differ significantly (post hoc Tukey tests).")
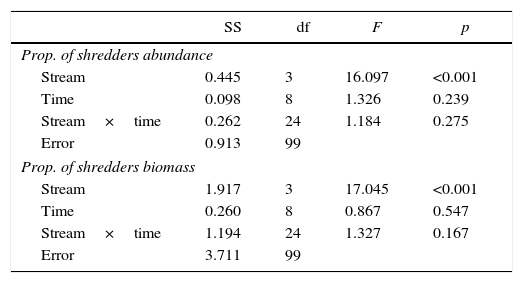
In total, 209 shredder organisms belonging to 6 different taxa (3 Trichoptera, 2 Plecoptera, and 1 Diptera) were found associated with leaves of M. chartacea (Table S2). The shredders composition found in Macuco and Banana differed from Norte and Luxemburgo (ANOSIM: R=0.34, p<0.01). In terms of abundance, the proportions of shredders in invertebrate assemblages were low throughout the experiment in Norte and Luxemburgo (<0.04±0.02). These values were constant and higher in Banana (0.10±0.04 to 0.27±0.15). However, the highest proportion of shredders was observed in Macuco after 150 days of incubation (0.42±0.18; Fig. S1A). In terms of biomass, the proportions of shredders were lower than 0.23±0.15 in Norte and Luxemburgo and ranged from 0 to 0.94±0.01 in Macuco and 0.12±0.05 to 0.54±0.24 in Banana (Fig. S1B). The natural logarithm of the proportions of shredder abundance and biomass differed among the studied streams only (Table 2), with higher values observed in Macuco and Banana than Norte and Luxemburgo.
Differences in the proportion of shredder abundance and biomass (ln[x+1] transformed data) in the invertebrate assemblages found in coarse mesh bags at different streams and experimental times tested by general linear models. SS=sum of squares. df=degrees of freedom.
| SS | df | F | p | |
|---|---|---|---|---|
| Prop. of shredders abundance | ||||
| Stream | 0.445 | 3 | 16.097 | <0.001 |
| Time | 0.098 | 8 | 1.326 | 0.239 |
| Stream×time | 0.262 | 24 | 1.184 | 0.275 |
| Error | 0.913 | 99 | ||
| Prop. of shredders biomass | ||||
| Stream | 1.917 | 3 | 17.045 | <0.001 |
| Time | 0.260 | 8 | 0.867 | 0.547 |
| Stream×time | 1.194 | 24 | 1.327 | 0.167 |
| Error | 3.711 | 99 | ||
Ecological gradients are ideal scenarios to assess how species interact and respond to changes in environmental conditions (Stevenson, 1997). Our results demonstrated that the conservation level in the riparian zones can influence leaf breakdown rates and the importance of invertebrate shredders in this process. Therefore, the evaluation of ecological processes in streams that have gradual changes in environmental conditions can be an alternative to better understanding of the effects that different disturbances have on aquatic communities and stream ecosystem functioning (Mittelbach et al., 2001).
The differences in the breakdown rates observed between coarse and fine mesh bags indicated that decomposer assemblages responded differently to the conservation level in the riparian zones. These results suggested that, at least in the studied streams, invertebrate shredders were more susceptible to disturbances in riparian zones. Of the total shredders sampled, 86% were found in the Macuco and Banana streams, and 2 taxa occurred only in these ecosystems. The low abundance of shredders in Norte and Luxemburgo probably resulted in slower breakdown rates. The decrease in the availability of food resources and habitat on the streambed might have influenced the distribution of shredders, such as that observed by Landeiro et al. (2010) in Amazonian streams with changes in the riparian vegetation. Furthermore, we noticed the deposition of fine sediment on leaves incubated in Norte and Luxemburgo, and this might have influenced the colonization by shredders (Niyogi et al., 2003). In this context, the abundance of shredders in leaf patches would be directly related to the conservation level in the riparian zones. Beyond the higher sensitivity of shredder taxa to stream environmental conditions (Masese et al., 2014), these invertebrates depend on leaf availability for food (Graça et al., 2001), habitat (Landeiro et al., 2010) and case-building (Rincón and Martínez, 2006).
Tropical streams differ in the abundance of shredders (Boyero et al., 2015), quality of available organic matter (Silva-Junior et al., 2014), water nutrient contents (Jinggut et al., 2012) and temperature (Boyero et al., 2014). These natural differences are mainly responsible for the variation in leaf breakdown rates across different regions (Boyero et al., 2015; Graça et al., 2015). The kc/kf ratio was already used to evaluate the importance of shredders in tropical streams (see Jinggut et al., 2012; Masese et al., 2014). In Brazil, values close to 1 were found in Cerrado streams, suggesting a low participation of shredders (Boyero et al., 2015). However, as far as we know, the kc/kf ratios have not been determined in streams with a higher abundance of shredders. The values observed in this study indicated a high importance of shredders in the decomposition process in more preserved streams, corroborating other studies that found high abundances of shredders in the Atlantic Forest (e.g., Oliveira and Nessimian, 2010). Gessner and Chauvet (2002) suggested that kc/kf ratios might vary substantially in streams that differed in the diversity of leaf patches, such as tropical streams.
Unlike invertebrate shredders, the activity of microbial decomposers, measured indirectly through fine mesh bags, only differed between the extremes of the four studied streams, i.e., Norte and Banana streams. However, the taxa composition and biomass of microorganisms that colonized the leaves might have differed among the streams (Martins et al., 2015). We expected that microbial activity would increase in more altered streams because changes in riparian vegetation can increase water temperature and nutrient content (Sridhar and Bärlocher, 1997). However, nitrogen and phosphorus concentrations were low, and the presence of riparian vegetation, even in disturbed streams, probably resulted in similar water temperatures (see Table S1). These results suggested that, in the studied streams, microbial activity was less affected by changes in the riparian zones, as was observed by Encalada et al. (2010) in streams with different riparian land uses.
Our results demonstrated that the conservation level in the riparian zones influenced the decomposition mediated by invertebrate shredders, supporting the use of this process as an effective indicator of stream functioning. Moreover, the study of leaf decomposition in streams with gradual differences in environmental conditions promoted a better evaluation of the effects that changes in riparian zones have on leaf breakdown rates and the importance of invertebrate shredders. Although breakdown rates in fine mesh bags had a lower response, the utilization of fine and coarse mesh bags in our study was important to determine the role of shredders in leaf decomposition. Finally, our results supported the notion of the importance of riparian zones conservation for the maintenance of aquatic communities and ecological processes that ensure the functioning of small forest streams, such those found in the Atlantic Forest.
Conflicts of interestThe authors declare no conflicts of interest.
We thank Carlos Burke for the logistical facilities, the Research Foundation of the State of Espírito Santo (FAPES; Proc. No. 61861480/2013) and the Brazilian National Research Council (CNPq; Proc. No. 449957/2014-4) for financial support to Marcelo Moretti and scholarships to the co-authors.
The following are the supplementary data to this article:

