







Climate conditions have a strong influence on the distribution of many natural communities; thus, the influence of global climate change may alter ecosystems. High-altitude wetlands in the Central Andes of Argentina (CAA) provide relevant ecosystem benefits and promote human activities. However, a degradation of Andean wetlands has been observed and documented in the last decades. Projecting the impacts of climate change on future distribution of wetlands is an essential subject in ecological studies. In this work, the present and future suitability for wetland systems in a pristine section of the CAA using spatial distribution modeling under low and high-emission scenarios are explored. The studied wetlands are strongly driven by bioclimatic variables such as mean annual temperature, precipitation, and its seasonality. Projections show that most of the currently occupied areas will modify under future climate conditions. Changes in temperature and precipitation patterns will decrease the potential suitability in low elevation areas for the next decades for the species inhabiting there, especially south of 29 °S. Additionally, future warmer climatic conditions, greater temperature variability, and reduction in precipitation would probably affect the snow cover and the available water supply which are key limiting factors for the distribution of Andean wetlands. This research is a contribution to understanding possible effects of climate change on high-altitude ecosystems. Mitigation measures for conservation of wetlands in CAA are immediately required to compensate for the impact of climate change under future environmental conditions.

High-altitude wetlands are one of the most important productive mountain ecosystems worldwide, being hotspots of biodiversity and relevant for the regional water cycle, and provide fundamental ecosystem benefits such as freshwater for many human settlements, livestock, and refuge for plants and animals inhabiting there. They are spread in mountain areas around the world and possess an especial ecological relevance due to their species richness and endemism (Mitsch et al., 2013; Rubio et al., 2017).
In the Central Andes of Argentina (CAA), wetlands are distributed at high altitudes and are characterized by extreme low temperatures, large thermal amplitude, snow cover during several months of the year, frozen soils (permafrost), and frequent strong winds (Martínez-Carretero and Ontivero, 2016; Mitsch et al., 2013). Andean wetlands are supplied by a continuous water flow that offers optimal conditions for associated plant communities (Carilla et al., 2018; Izquierdo et al., 2022; Ruthsatz et al., 2020). They provide numerous local benefits such as flooding control, water supply and regulation, aquifer recharge, nutrient recycling, erosion control, terrestrial carbon storage, fixation, and soil formation (Mitsch et al., 2013). Moreover, they are important for regional climate regulation with a strong influence in the water cycle and rain production (Bullock and Acreman, 2003). The presence and ecological importance of wetlands is greater in places where water resources are scarce as the case of arid and semi-arid ecosystems in southern South America (Martínez-Carretero and Ontivero, 2016). The geographic position of high-altitude wetlands makes them highly vulnerable to changes in temperature and precipitation patterns (Cuesta et al., 2023; Ontivero, 2015).
Climate change is the major threat to mountain ecosystems at a global scale (Huggel et al., 2019; Pauli and Halloy, 2019) and is expected to become the main driver of biodiversity and ecosystem degradation in the next decades (Malhi et al., 2020). Currently, the hydrological cycle in the Andean region is seriously affected by the increase of temperature causing glacier retreat and alterations in the precipitation patterns, thus placing the Andean wetlands at a high vulnerability level (Bradley et al., 2006; Hugonnet et al., 2021; Rivera et al., 2021). High-altitude wetlands are strongly sensitive to global warming because they are mainly handled by low temperatures throughout the year. The degradation of these fragile ecosystems has been documented at a global scale (Dangles et al., 2017; Durigan et al., 2022). Thus, they are seriously threatened all over the world and a call to avoid a declining availability of fresh water has been made (Finlayson et al., 2019). A modification and loss of these areas is projected for the next decades with a consequent negative impact in natural habitats and a high risk of disappearance of many endemic species (Cuesta et al., 2023; Sala et al., 2000). Studies affirm that the mean temperature in the CCA will increase significantly under future climate conditions, particularly under high emissions scenarios (Rivera et al., 2020). Therefore, it is expected that climate changes may produce serious structural and functional modifications of wetland dynamics with the limitation in the provision of ecosystem services (Cuesta et al., 2023; Salimi et al., 2021). In the CAA, the increase in the aridization processes of high-altitude wetlands in natural protected areas was highlighted (Ontivero, 2015; Martínez-Carretero and Ontivero 2016). Species divergent responses and diversity losses in Andean biomes have been projected under future climate scenarios (Tovar et al., 2022). However, the quantification of impacts and potential distribution of high-altitude wetlands under future climate conditions are still unclear. For this reason, awareness of their present suitable distribution areas in the CAA related to climate variables is essential for understanding the future perspectives of these delicate mountain ecosystems (Carilla et al., 2018; Dangles et al., 2017). Knowing the current distribution area occupied by high-altitude wetlands in the CAA is relevant for comprehending the structure and functioning of these ecosystems under global climate change. Here, the principal hypothesis deals with a change of suitable areas for high-altitude wetlands in CAA under future climate conditions. Therefore, in this research, the present and future distribution of high-altitude wetlands in two natural protected areas from the CAA were explored. Projected changes under future climate conditions using two greenhouse gasses emissions scenarios were analyzed, and the effects of prospective impacts on their structure and functioning were discussed.
The analysis of climate projections and the response of biological processes help to understand future impacts on natural systems (Parmesan et al., 2000). Moreover, the knowledge about the potential and future distribution of Andean wetlands will contribute to the implementation of conservation practices, placing information available for the decision makers (Crego et al., 2014; Ontivero et al., 2022). Thus, the effectiveness of management practices demands a better knowledge of how these ecosystems will change and adapt under future variations in regional temperature and precipitation patterns.
MethodsStudy areaHigh-altitude wetlands in the CAA are mainly supplied by groundwater along the Puna and the Altoandina phytogeographic provinces (Martínez-Carretero and Ontivero, 2016). The study area comprises two adjacent natural protected areas in the CAA (Fig. 1). The San Guillermo protected area (28º 27' S, 69º 32' W) is located in the northwest of San Juan province, at an altitude of 1,700–5,000 m. On the other hand, the Laguna Brava protected area (28º 28' S, 69º 10' W) is located in the southwest of La Rioja province and characterized by a system of shallow lagoons of saline and hypersaline water at altitudes of around 2,500–4,500 m (Wurstten et al., 2014). The protected areas encompass nearly 18,000 km2 and have become exposed, in many sections, to anthropogenic disturbances such as mining, livestock, and tourism (Didier et al., 2009; Silvani et al., 2017). Due to their regional importance and fragility, they were incorporated into conservation programs for protecting wetlands and their associated diversity. The protected area of San Guillermo was declared as Reserve of the Biosphere by UNESCO in 1980 while Laguna Brava was incorporated into the Ramsar Convention in 2003. These areas protect and preserve a relevant biodiversity associated with the local wetlands and plant communities, promoting the presence and reproduction of flamingos and camelids.

The regional climate conditions are strongly influenced by the pronounced altitudinal gradient and the complex topography. The area is characterized by a wide thermal amplitude, with mean monthly temperatures ranging from 9 °C in summer to −7 °C in winter, strong cross-barrier winds, and most of the scarce precipitation (approximately 100 mm per year) falling as rain during summer months, with occasional snowfall during the winter season (Wurstten et al., 2014). The vegetation cover forms an extensive meadow, distributed in an altitudinal range from 3,800 to 4500 m with dominance of microthermal floristic components (Ontivero et al., 2022). It is composed of an association of grass steppes and shrubby species mostly in the area of the slopes with wetland ecosystems distributed in the lowlands.
Occurrence dataA database of georeferenced occurrences of the high-altitude wetlands was constructed from remote sensing using Sentinel 2B images from 2019 at a scale 1:50.000. The occurrences were selected on the images following the wetlands classification based on the local geomorphology and hydrology proposed by Ontivero et al. (2022). High-altitude wetlands are groundwater dependent systems in a close association with vegetation communities sharing high climate and environment affinities (Méndez, 2007). Recently, distribution models have been used to analyze wetlands as entities (Raney and Leopold, 2018). Therefore, high-altitude wetlands placed in the studied area were analyzed as entities, recording their coordinates for conducting the present study. Additionally, many of the identified wetlands were visited during different field trips to the area and their conservation status was discussed by Ontivero (2015). A total of 310 data points were recorded for both natural protected areas in zones with reduced mining activities and grazing (Fig. 1).
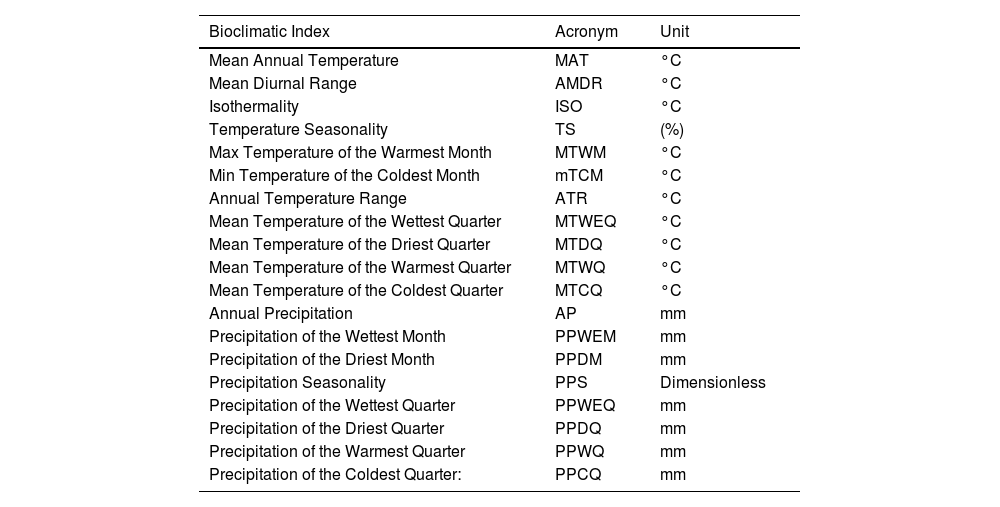
Climate variable selectionAn initial set of 19 bioclimatic variables in spatial resolution of 30 arc s (∼1 km2) from the CHELSA v2.1 database (Karger et al., 2020) were used to characterize the distribution of high-altitude wetlands in the study area. This database has higher accuracy in mountain regions than other precipitation and temperature datasets, given that it better represents orographic precipitation effects (Bobrowski et al., 2021). The bioclimatic variables (Table 1) were derived from monthly temperature (maximum and minimum) and precipitation data that represent mean annual values, seasonality, and extreme conditions (O’Donnell and Ignizio, 2012). To avoid collinearity and select the most relevant bioclimatic parameters, the total of variables was reduced using the Spearman’s correlation, with a threshold of |r| = 0.80. Correlated variables were removed based on their highest contribution.
Biolimatic variables used in the modeling analysis.
| Bioclimatic Index | Acronym | Unit |
|---|---|---|
| Mean Annual Temperature | MAT | °C |
| Mean Diurnal Range | AMDR | °C |
| Isothermality | ISO | °C |
| Temperature Seasonality | TS | (%) |
| Max Temperature of the Warmest Month | MTWM | °C |
| Min Temperature of the Coldest Month | mTCM | °C |
| Annual Temperature Range | ATR | °C |
| Mean Temperature of the Wettest Quarter | MTWEQ | °C |
| Mean Temperature of the Driest Quarter | MTDQ | °C |
| Mean Temperature of the Warmest Quarter | MTWQ | °C |
| Mean Temperature of the Coldest Quarter | MTCQ | °C |
| Annual Precipitation | AP | mm |
| Precipitation of the Wettest Month | PPWEM | mm |
| Precipitation of the Driest Month | PPDM | mm |
| Precipitation Seasonality | PPS | Dimensionless |
| Precipitation of the Wettest Quarter | PPWEQ | mm |
| Precipitation of the Driest Quarter | PPDQ | mm |
| Precipitation of the Warmest Quarter | PPWQ | mm |
| Precipitation of the Coldest Quarter: | PPCQ | mm |
The potential distribution model was conducted using Wallace EcoMod 2.1 (Kass et al., 2018) in R (R Core Team, 2021) and the algorithm implemented in the maximum entropy (MaxEnt) method (Phillips, 2021). As the program works with georeferenced data points; the 310 sites corresponding to the occurrence data were added into the program for the analysis. A spatial filtering (spThin R package) at a minimum distance of 1 km among occurrence data was applied to reduce clustering caused by sampling bias (Aiello-Lammens et al., 2015). The background extent was set to minimum convex polygon. The buffer distance was 0.1 degrees and 10,000 background points were sampled. Block spatial partitioning was used and occurrence data were partitioned in four groups based on spatial clustering of occurrence points (Brown, 2014). To avoid model overfitting, regularization multiplier values ranging from 1 to 5 (with 0.5 steps) were set. A combination of four feature classes: linear (L), quadratic (Q), product (P), and hinge (H) were tested. The model evaluation was run through the ENMeval R package (Kass et al., 2021) and the selection was made considering the lowest Corrected Akaike Information Criterion (AICc) that corrects for sample size (Symonds and Moussalli, 2011; Warren and Seifert, 2011). The tenth percentile training presence threshold (10%TP) to provide a more comprehensive evaluation and quantification of the projected changes was applied. The presentation of the results in continuous grids were converted to binary grids (presence/absence) to discriminate between suitable and unsuitable areas. Although the threshold selection is still controversial, the 10%TP results are one of the most used and less restrictive, offering a good performance (Escalante et al., 2013).
Climate projectionsTo evaluate the current and future distributions of the wetlands the selected set of bioclimatic variables available in the CHELSA v2.1 dataset were used. Bioclimatic layers were constructed using five general circulation models (GCMs) from the Coupled Model Intercomparison Project Phase 6 (CMIP6) (Eyring et al., 2016), in particular those selected for the Intersectoral Impact Model Intercomparison Project version 3b (ISIMIP3b, Lange, 2019).
The models from which data were taken are: GFDL-ESM4, IPSL-CM6A-LR, MPI-ESM1-2-HR, MRI-ESM2-0, and UKESM1-0-LL, for a spatial resolution of 30-arc-second. Climatic data of the selected bioclimatic variables for a reference period (1981–2010) and for the mid-term time horizon (2041–2070) using the SSP1-2.6 and SSP5-8.5 scenarios as a forcing pathway. The SSP1-2.6 scenario will produce a multi-model mean of significantly less than 2 °C warming by 2100, and therefore can support analyses of this policy goal (O’Neill et al., 2016). The SSP5-8.5 represents a high fossil-fuel development throughout the 21st century, with a maximum projected global temperature change of almost 5 °C for 2100 (Meinshausen et al., 2020).
A multi-model ensemble considering the average of the five GCMs was obtained in order to reduce the uncertainty related to the model biases. This allowed us to obtain a single climatology for the reference period that was compared to the future climate in order to obtain differences among the bioclimatic indicators and, thus, the potential distribution of the high-altitude wetlands in the CAA.
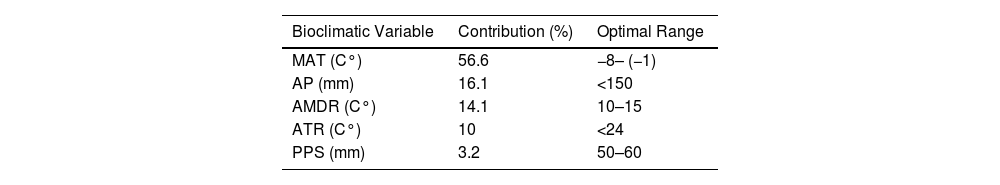
ResultsClimate variables and model selectionThe set of uncorrelated and best predictor variables after the distribution model analysis were: MAT, AMDR, ATR, AP, and PPS (Table 2). The model with the highest performance (lowest AICc = 4852.98) identified the optimal beta-multiplier and set of the most relevant bioclimatic variables, which were used to project habitat suitability of wetland ecosystems in the CAA both the reference and the future periods. According to the Maxent models, the lowest AICc scores and highest area under curve (AUC) for the training dataset were assigned to the model with a beta-multiplayer of LQ4. The calculated value of the AUC was 0.72 which indicates a good model performance. The climatic response curves obtained from the analysis indicated the optimal values of the five bioclimatic variables, which model the current climate conditions for the studied wetlands (Table 2).
Climate variable contribution and optimal range values for the five uncorrelated and most relevant variables.
| Bioclimatic Variable | Contribution (%) | Optimal Range |
|---|---|---|
| MAT (C°) | 56.6 | −8– (−1) |
| AP (mm) | 16.1 | <150 |
| AMDR (C°) | 14.1 | 10–15 |
| ATR (C°) | 10 | <24 |
| PPS (mm) | 3.2 | 50–60 |
MAT: mean annual temperature; AP: annual precipitation; AMDR: annual mean diurnal range; ATR: annual temperature range; PPS: precipitation seasonality.
Considering the selected bioclimatic indices, a suitability map showing the potential distribution of high-altitude wetlands was obtained. Areas with higher suitability values are mainly located in an altitudinal range from 3,000 to 4,500 m, while those areas with low suitability (0–0.2) are located in the southeastern part of the San Guillermo protected area which corresponds to low altitudinal range low suitability areas are also observed in the western edge of this protected area, a factor that is linked to the presence of high-elevation areas with altitudes above 4,500 m. On the other hand, Laguna Brava protected area shows a high suitability for wetlands in most of its area, especially in the central part of the reserve, i.e., with altitudes ranging from 3,500 to 4,500 m, where the highest suitability values were obtained (Fig. 2).
Future changes in Andean wetlands.")
The resulting suitability map for high-altitude wetlands in the CAA using the projected bioclimatic indices for the SSP1-2.6 scenario shows a general reduction in the suitable areas for the period 2041-2070 compared to the reference period, particularly over the San Guillermo area (Fig. 3A). Much of the central and southernmost parts of the San Guillermo area has a projected reduction in suitability compared to the present, mainly for the areas located at elevation lower than 4,000 m. The projected reductions over the Laguna Brava protected area are located in the western portion of the region (Fig. 3B), where a large number of wetlands are placed (Fig. 1). The projected change in suitability shows higher percentages (i.e. more suitable future conditions for high-altitude wetlands) in those zones constrained to elevation values higher than 4,000 m (Figs. 1 and 3B).
 Future suitability for high-altitude wetlands in the CAA for the period 2041–2070, based on the SSP1-2.6 emissions scenario. (B) Projected suitability change for the distribution of wetlands in the study area for the same period and emissions scenario.")
Based on the SSP5-8.5 scenario, results show a similar spatial pattern in terms of positive and negative projected changes in suitable areas for the period 2041–2070 (Fig. 4A–B). The most suitable areas are restricted to the northern part of the study area, mainly along the Laguna Brava protected area and the northeastern portion of San Guillermo protected area. Areas with low suitability (0–0.2) are projected mostly for the western and southeastern regions of San Guillermo protected area (Fig. 4A), which also correspond to low suitability areas based on the reference period (Fig. 2). The projected change in suitability shows higher percentages in the highest zones of the study area (Fig. 4B). This becomes more evident for the eastern section of Laguna Brava protected area, and the northern portion of San Guillermo protected area. The regions with elevation higher than 4,000 exhibit high percentage values, which means a projected positive change in suitability. Most of the San Guillermo area shows a negative change in suitability, particularly for those areas with altitude values lower than 4,000 m. The area with the highest density of sites in the western portion of Laguna Brava, also shows a projected decrease in suitability.
 Future suitability for high-altitude wetlands in the CAA for the period 2041–2070, based on the SSP5-8.5 emissions scenario. (B) Projected suitability change for the distribution of wetlands in the study area for the same period and emissions scenarios.")
Projected climate change using a threshold confirms the placement of future optimal regions in the northern part of the studied area and deterioration towards the southern part. The projected changes in the distribution of high-altitude wetlands display an elevation dependence (Fig. 5A–C). The suitability for the studied wetlands was discriminated for the reference as well as for both future periods by the altitudinal bands (Fig. 6). The projected decrease in suitability can be linked to elevations below the 4,200 m a.s.l. with abrupt changes projected for the altitudinal range of 3,000 to 3,400 m a.s.l., and 3,600 to 3,800 m a.s.l. It is observed a projected increase in suitability in those areas located at elevations higher than 4,200 m a.s.l. The most notorious difference is shown between the projected future scenarios and the reference period for the regions located above 4,400 m a.s.l. It must be noted that the figure shows an optimum present suitability range for wetlands placed between 3,600–4,000 m a.s.l., sharply declining to elevations higher than 4,400 m a.s.l. (Fig. 6).
. (A) Reference period (1981–2010). (B) Period 2041–2070, based on the SSP1-2.6 emissions scenario. (C) Period 2041–2070, based on the SSP5-8.5 emissions scenario.")

High-altitude wetlands in CAA are a combination of evergreen meadows in a hydromorphic soil supply by groundwater and running on a permanent water flow (Martínez-Carretero and Ontivero, 2016; Ontivero, 2015). Therefore, the distribution and presence of these ecosystems are in association with the regional available water resources and vegetations. Plant communities associated with wetlands in CAA are frequently distributed in altitudinal zonations with vegetation records rare or scarce at elevations between 4,200–4,300 m a.s.l. and practically absent for elevations over that range (Méndez, 2004; Ontivero, 2015).
Recent studies showed that changes in temperature and precipitation have disturbed the habitat of the Andean vegetation, limiting its distribution to small favorable refuges (Ruthsatz et al., 2020) with the consequent loss of native species (Schuchardt et al., 2023). In the studied protected areas, an increase of dried wetlands and habitat modifications with loss plant coverage and diversity, plus soil deterioration, have been observed during the last years. The status classification and analysis proposed by Ontivero (2015) for six wetlands in the CAA indicated that at least four were in a regular and intermediate conservation status with a high aridization percentage. In the San Guillermo natural protected area, high suitability areas are placed at an elevation range from 3,000 to 4,000 m. Low suitability ranges were in concordance with the lowlands (<3,000 m) placed in the southeastern part of the area, where the water supply for the development of wetlands is scant. Additionally, areas with low suitability are spread in the western part of the CAA at elevations above 4,500 m, a region characterized by rock glaciers and permafrost, with a geological setup that precludes the abundance of wetlands and vegetation. On the other hand, Laguna Brava natural protected area has a more equal distribution of highlands and suitability ranges (Fig. 2).
The present distribution of high-altitude wetlands in the CAA is strongly influenced by bioclimatic variables linked to temperature and precipitation (Fig. 2; Table 2). Seasonal precipitation is regularly stored as snowpack, playing a major role in determining plant composition. Precipitation either as rainfall or snow exerts a large influence in shaping wetlands in the Andes (Cooper et al., 2019; Feeley et al., 2020). Temperature combined with the spatial and temporal pattern of seasonal precipitation are relevant in determining the growing season in mountain ecosystems where plants spend a long dormancy due to frozen days and snow cover (Cavieres and Sierra-Almeida, 2018; Wipf and Rixen, 2010). In high-altitude ecosystems, snowmelt defines the onset and length of the growing warm season as well as the available water and nutrients that influence soil moisture and nutrient status for plants (Keller et al., 2005; Körner and Hiltbrunner, 2021). The regional temperature interacts with many components of the hydrological cycle, especially in determining the duration of the snowpack, a factor that affects the cover, diversity, and productivity of high-altitude plant communities (Wipf and Rixen, 2010). The MAT and ATR are also linked to water supply from glaciers, mainly in periods when snow has been completely melted (Crespo et al., 2020). Thermal daily and seasonal range have a relevant role in many physiological processes as flowering and seed production for high-altitude plants (Hülber et al., 2010). This relationship at daily and seasonal scale highlights the role of the temperature in the suitability of wetlands probably due to the modulation of water availability. In this sense, the warming recorded during the last decades in the Alps has caused plants with low thermal requirements to retreat towards more elevated areas (Körner and Hiltbrunner, 2021). In South America, several plant communities have experienced a thermophilization, positively correlated with warming rates (Feeley et al., 2020). Warming effect has caused a long-term reduction in glaciers along the CAA, a situation that has been exacerbated during the last decade (Dussaillant et al., 2019), along with the occurrence of extreme drought conditions that affected regional water availability (Rivera et al., 2021). Glacier retreat has a direct impact on wetlands (Bradley et al., 2006), and the sustained warming is causing a decline of permafrost (Rangecroft et al., 2016), a cryosphere component that plays a significant role as a water source in the study area (Esper-Angillieri, 2017).
Perspectives of Andean wetlands in the forthcoming decades under climate change scenariosAn increase in warming is projected for the next decades with serious effects for the highest altitudes of the CAA (Rivera et al., 2020), which implies an increase of regional temperatures from 1 °C to 4 °C by the end of the 21st century (Almazroui et al., 2021). Moreover, a reduction in the precipitation, runoff, and snow cover are projected for the rest of the century, even when the most optimistic emission scenarios are considered (Rivera et al., 2020; Almazroui et al., 2021). Analyzing the projected changes according to the selected bioclimatic indices for the two greenhouse gasses emissions scenarios, a negative change in the suitability for high-altitude wetlands is projected, particularly towards the south of 29 °S and the lower elevation areas (Figs. 3A and 4A). Projections for wetlands in the southernmost Andes of Patagonia indicated that suitable areas will decline under future climate conditions (Crego et al., 2014). A warmer climate will produce changes in life conditions for high mountain ecosystems at very small scales, mainly as a consequence of the reduction in snow duration, with species moving towards more optimal thermal habitats (Körner and Hiltbrunner, 2021). In this sense, species with a cold period required for seed germination may be endangered in the future (Cavieres and Sierra-Almeida, 2018).
Suitable areas in San Guillermo and Laguna Brava protected areas will be restricted at high-altitudes in the CAA (Figs. 3A and 4A) with a probable reduction of vegetation cover and composition. Recently, changes in vegetation cover and species richness for the CAAs in the north of Argentina were recorded with the migration of species towards wider thermal niches (Cuesta et al., 2023). Environmental changes, linked to regional climate variability, were detected in wetlands from the northwestern part of Argentina and southeastern Bolivia. These studies recorded a severe aridization trend in the Andean wetlands and a decrease in the plant productivity several decades ago (Carilla et al., 2013; Morales et al., 2015). Vegetation communities will probably reshape highland ecosystems in the next decades with different distribution patterns, abundances, and specific composition (Feeley et al., 2020).
The reduction of wetlands is the result of soil dryness and aridization with the loss of organic matter and the disappearance of the plant communities that formed these ecosystems. Therefore, the deterioration of wetlands in response to temperature encourages the loss of biodiversity. Additionally, an early snowmelt under future climate would imply snow-free mountain areas during prolonged periods that could promote the invasion by species potentially more efficient in flowering than natives (Keller et al., 2005; Martínez-Carretero and Ontivero, 2016; Schuchardt et al., 2023). According to the projected models, the most suitable areas with a greater presence of wetlands in the future will be restricted to altitudes of more than 4,200 m a.s.l. (Figs. 5A–C and 6). These areas will keep adequate climate conditions for high-altitude wetlands and probably for vegetation migration.
However, there are permanently frozen soils and pronounced slopes with intense cryogenic processes and soil mobilization in the CAA at altitudes above 4,600 m a.s.l. that makes the rooting of plants difficult (Méndez, 2007).
Even though the future changes in suitability indicate better climate conditions for the north part of the studied area in the higher altitudes, these ecosystems may experience a reduction with a high risk of disappearing in the next decades (Figs. 3A, 4A, and 5A–C). In the CAA, most of the wetlands face limited provincial or national environmental regulations in addition to human impacts such as mining, tourism or livestock. There are a few studies which attempt to define local parameters for evaluating their present and future conditions. Monitoring wetlands is an essential task that should be considered for preservation of these natural areas (Carilla et al., 2013; Cuesta et al., 2023; Ontivero, 2015).
The conservation of these sensitive high-mountain ecosystems will ensure that water resources originating in the Andes will be sustained in the future under the increased global warming conditions (Finlayson et al., 2019).
ConclusionsClimate change is a global concern related to human activities. Drastic consequences on the Earth’s ecosystems will rapidly increase as temperature continues rising.
For the Andean wetlands, placed in two natural protected areas from the CAA, changes in their distribution areas are projected for the period 2041–2070 under a low and a high-emissions scenario relative to the 1981–2010 period. Undoubtedly, the present distribution of high-altitude wetlands in the CAA will be modified under future climate change. The projected increase in temperature, together with a decrease in precipitation will have a direct effect on the suitable conditions, influencing the current distribution of Andean wetlands. Most of the negative changes in the distribution of wetlands will occur at altitudes below 4,200 m. The projected changes, considering the most relevant bioclimatic indices, will affect the future distribution of high-altitude wetlands, probably related to the reduction in water supply at lower elevations. These changes probably affect the plant communities, diversity, and their distribution in Andean wetlands following the reduction of the snow cover and cryosphere components such as glaciers and permafrost. High optimum areas will be placed at altitudes of more than 4,200 m. a.s.l. However, the vegetation and the associated life forms in areas of more than 4,500 m a.s.l. are currently restricted to the topography. It means a narrow range of possibilities for the existence of wetlands and green lands in the long term and a high risk of the disappearance of many endemic and native plants. Understanding their present conditions and any potentially threatening such as global climate change are essential to take action. Management practices for preserving these ecosystems are necessary in the short term. Our study represents the first step to explore the present and future distribution suitability of high-altitude wetland systems in CAA as regional entities under changing climate conditions. The intrinsic value of high Andean wetlands, the provision of ecosystem benefits, and the climate threats have led to increased awareness about the necessity of actions to protect and preserve these natural environments.
Conflict of interestThe authors declare not any conflict of interest.
Financial supportThis research was supported by the National Agency for Scientific and Technological Promotion (ANPCyT) and the University Juan Agustín Maza (Grant No. PICTO-UUMM-2019-00004).

