



Extensive invasion of the Campos grasslands of southern Brazil by the South African grass, Eragrostis plana, is a serious conservation problem due to its low palatability and ability to quickly spread. Here, we experimentally evaluated the effects of grazing management and initial disturbance on invasibility of native grassland community by E. plana. The experiment, a split-plot complete block design with three replications, was established in 2004 in non-invaded native grassland. Grazing management treatments were assessed in main plots (continuous grazing, rotational grazing, and grazing exclusion). The initial disturbance treatments (light grazing, heavy grazing, and heavy grazing plus soil scarification) were applied in subplots before the sowing of standard amounts of E. plana seeds. The initial disturbances heavy grazing and heavy grazing plus soil scarification simulated the introduction of exotic forage species. We monitor plant species composition and diversity along six years after the experiment starting. The effects of grazing management and initial disturbance on community invasibility were significant and showed interaction. Heavy grazing plus soil scarification increased the rate of E. plana invasion and produced more pronounced changes in species composition than the other treatments. Plots under grazing exclusion, rotational or continuous grazing without soil scarification had low levels of invasion (<10% cover). But exclusion also changed community composition and decreased diversity, while rotational and continuous grazing maintained community structure. The invasion can be largely prevented on undisturbed grassland by employing either rotational or continuous grazing management. If the livestock producers want to introduce another forage species in their natural grasslands and, at the same time, do not run the risk of opening space for E. plana should avoid the use of soil scarification.
Biological invasions by exotic species around the world have increased during the recent decades perhaps due to the increase of global trade and the opening of new markets (Pyšek et al., 2010). Invasion by Eragrostis plana (lovegrass), a South African native species locally known as capim-annoni, has been causing damage to native grasslands, affecting the grasslands since community composition, structure, diversity, forage quality, and social systems in southern Brazil (Medeiros et al., 2009). E. plana is widely spread in this region mainly along roadsides and in compacted soil areas. This species also has some features that give it competitive advantages in relation to native species as the high production of small viable seeds presenting dormancy (Medeiros et al., 2014), development even under low levels of soil nutrients due to its photosynthetic efficiency and anatomical features, high levels of lignin, which makes it unpalatable for cattle, and the ability to store starch on roots (Favaretto et al., 2015a). By 2004, it was estimated that over one million hectares had been already invaded by E. plana mainly in the native grasslands of southern Brazil (Medeiros and Focht, 2007). However, bioclimatic modelling of the habitat suitability in South America has predicted that the occupied region may greatly increase (Barbosa et al., 2013).
The southern Brazilian Grasslands (Campos) are native ecosystems holding high species richness (Boldrini, 2009), where disturbances by grazing and/or fire are essential under present climatic conditions to maintain the physiognomy and plant diversity (Overbeck et al., 2007). These ecosystems feature a long history of coevolution between plant species and large grazers until the extinction of the latter by the end of the Pleistocene (Cingolani et al., 2005; MacFadden, 2000). Thereafter anthropogenic fire played a key role in maintaining open vegetation until the introduction of cattle and horses at the 18th century (Behling et al., 2007; Müller et al., 2012). Since 1970, these grasslands have been strongly affected by land use changes leading to increased conversion into annual croplands (mainly soybean and rice), forest plantations, planted pastures with exotic species (Overbeck et al., 2007) and invasion by E. plana (Guido et al., 2016; Medeiros et al., 2009).
Livestock producers manage natural grasslands through different ways, mostly manipulating the grazing intensity and introducing exotic species in order to increase the forage productivity. The intensity of grazing is recognized as a regulator of the competition processes in grasslands plant community, altering vegetation structure parameters such as species richness and evenness. In moist pastures, such as those found in the southern Campos, it is common to find intensely grazed areas with greater richness of species, benefiting forbs and grasses species with physiological and morphological adaptations to survive under intense grazing (Cingolani et al., 2005). At lower intensities, there is a tendency to decrease plant richness and predominate species with greater competitive potential, such as caespitose grasses, greatly reducing species evenness (Hendon and Briske, 2002; Škornik et al., 2010). Therefore, by manipulating the intensity of grazing, livestock producers influence local resource availability and the balance of structuring community processes (Davis et al., 2000). Other management strategy used by livestock producers is the introducing of exotic forage species aiming the increase of forage amount in winter when native grassland species are less productive. This strategy is often accompanied by soil disturbance or heavy grazing, which breaks with the coverage structure of the communities, altering light availability, exposing bare soil and nutrients, and weakening local species (Hobbs and Huenneke, 1992). Therefore, both management actions modify the resources availability and the balance of competition and facilitation interactions (Bruno et al., 2003) and thus they can change the invasibility of the grassland communities (Hobbs and Huenneke, 1992).
Disturbances largely influence community assembly and invasibility of ecosystems (Davis et al., 2000; Colautti et al., 2006). In general, invasion processes are determined by several factors that co-vary in space and time, including climate (Parepa et al., 2013), community structure (Tilman, 2004; Fargione and Tilman, 2005), disturbances (sensuHobbs and Huenneke, 1992) in different scales (Stohlgren et al., 2006), resource availability (Davis et al., 2000), propagule pressure (Colautti et al., 2006) and related with ecosystem processes (Moles et al., 2012). Endogenous community factors are considered to define the resistance of communities to invasion and they may be biotic or abiotic. Biotic resistance is the reduction in the success of the invasion caused by competition or disease (Levine et al., 2004; Kuebbing and Nuñez, 2015), while abiotic resistance is when the invader does not tolerate a given habitat condition (Davis et al., 2000), such as the restriction of a given species to a climatic condition. Yet, since abiotic conditions influence the biota and vice versa, the interaction of both sources of resistance should be considered when assessing invasion processes.
Here we report results of a six-year experiment assessing the effects of grazing management (rotational grazing, continuous grazing and grazing exclusion) combined with different types of initial disturbance (two levels of grazing intensity and soil scarification) on plant community dynamics and invasion by E. plana during this period. E. plana was sowed at high seed density. We assessed species composition, richness, evenness and diversity of plant communities during six years after the starting of the experiment, as well as the amount of E. plana coverage. We hypothesized that the treatments that simulate conditions for forage species introduction, i.e. those that first alter the community structure with heavy grazing with or without soil scarification, will result in invasion by E. plana contrasting with the treatment of light grazing. The greater change in the initial conditions (i.e. heavy grazing with soil scarification), more intensive will be the invasion. We even predict that such initial changes will influence community structure parameters, leading to differences between treatments concerning diversity, evenness, and species composition trajectories along the observed period. Moreover, we expected changes in community structure due to differences on simulations of grazing management. Rotational regime would allow a plant community with greater resistance to invasion by E. plana due to higher diversity and evenness, while grazing exclusion will promote unevenness communities, with low invasion rates, but with a very distinct trajectory characterized by low richness and tussock grasses dominance.
MethodsStudy areaThe experiment was established in a native grassland paddock (31°5′53.83″S, 54°57′37.44″W) with 8100 square metres, located at Estância Upacaraí farm, municipality of Dom Pedrito, Rio Grande do Sul, Brazil (see Focht and Borges de Medeiros, 2012 for more details). The area has been grazed by cattle and horses since the 17th century, when grazers were introduced in the region. E. plana was absent in the experimental site, but was present in nearby paddocks and roadsides. The climate type is Cfa (Köppen classification) with 1430mm annual mean precipitation and frequent water deficits during summer (Alvares et al., 2013). The annual mean temperature was 20.2°C, while the average temperature of the warmest month and the coldest month was 24°C and 12.4°C, respectively. The soil corresponds to Chernozems (IUSS Working Group, 2015).
Experimental designThe experiment was a split-plot design with three complete 30×90m blocks (Appendix S1). Three grazing treatments were applied to the main plots (30×30m): (1) continuous grazing by cattle (Bos taurus), (2) rotational grazing and (3) grazing exclusion. Further, before the sowing of E. plana, three disturbance treatments were applied once to the subplots (30×10m) within each main plot in July 2004: (1) light grazing that left an aboveground biomass residue of ca. 10cm height (LG); (2) heavy grazing that left and aboveground residue of ca. 5cm height (HG); and (3) heavy grazing that left and aboveground residue of ca. 5cm height plus soil scarification by rotatory tilling to 10cm depth (HG+SC). Treatments (HG) and (HG+SC) also included the sowing of a seed mixture with Lolium multiflorum (25kgha−1), Lotus corniculatus (6kgha−1), and Trifolium repens (2kgha−1), which are forage species commonly used in winter pastures regionally.
The experiment was a split-plot design with three complete 30×90m blocks (Appendix S1). Three grazing treatments were applied to the main plots (30×30m): (1) continuous grazing by cattle (Bos taurus), (2) rotational grazing and (3) grazing exclusion. Further, before the sowing of E. plana, three disturbance treatments were applied once to the subplots (30×10m) within each main plot in July 2004: (1) light grazing that left an aboveground biomass residue of ca. 10cm height (LG); (2) heavy grazing that left and aboveground residue of ca. 5cm height (HG); and (3) heavy grazing that left and aboveground residue of ca. 5cm height plus soil scarification by rotatory tilling to 10cm depth (HG+SC). Treatments (HG) and (HG+SC) also included the sowing of a seed mixture with Lolium multiflorum (25kgha−1), Lotus corniculatus (6kgha−1), and Trifolium repens (2kgha−1), which are forage species commonly used in winter pastures regionally.
The continuous grazing treatment was applied according to the local farm management regime by allowing free cattle access into the assigned main plots. The mean stocking rate in this treatment was 0.7 animalsha−1 (280kg live weight of cattleha−1), aiming at maintaining the sward height at ca. 5cm (recorded mean was 6.14±1.6cm). The main plots assigned to the rotational grazing treatment were fenced and temporarily grazed about eight times per year by cows that were fasted during the previous night. Cows were maintained in each plot until sward height was reduced to ca. 10cm (recorded mean was 11.37±3.7cm). This rotational grazing treatment was maintained until mid 2007 (34 months after the beginning of the experiment). Due to logistic problems in maintaining the rotational system, rotationally grazed plots were continuously grazed from mid 2007 to early 2011. Thus, this treatment was named rotational-continuous in the analyses that included data from the 2010 survey (see further details below). For the grazing exclusion treatment, plots were just fenced and no cattle access was allowed until early 2011. For details about the treatments see Focht and Borges de Medeiros (2012).
The grazing regimes and disturbance levels resulted in nine treatment combinations. The subplots were further divided into three 2×2m sub-subplots, on which measurements were done. In these units, immediately after initial disturbances were applied, we sowed E. plana in all treatments in July 2004 and in July 2005 (1539 seeds m−2 year−1), to standardize propagule pressure. The weight of 1000 seeds of E. plana was 0.23g with 95% germination (Medeiros et al., 2014).
Data collection and analysesPlant species cover was monitored in a 0.5×0.5m permanent quadrat positioned in the centre of each 2×2m unit. We used the Braun-Blanquet scale for visual estimation of cover-abundance (Mueller-Dombois and Ellenberg, 1974), which was then transformed into percentages as follows: +: 1%, 1: 2.5%, 2: 15%, 3: 37%, 4: 62% and 5: 87%. For the analyses, we considered the average of the three units in each subplot. Data collection started in May 2004 (prior to treatment implementation), October 2004 (four months after), April 2005 (10 months), November 2005 (17 months), May 2006 (23 months), November 2006 (29 months), April 2007 (34 months) and November 2010 (77 months). In order to sample the representative summer and spring species composition, we conducted the surveys twice every year.
We calculated the rate of increase of E. plana cover by taking the difference in E. plana cover for each two subsequent surveys divided by the time elapsed in months. Finally, we calculated the mean values of cover increment rate between month 0 and 34. For this we only considered the surveys conducted between the months 0 to 34, since the rotational grazing was thereafter changed.
For assessing the effects of the experimental factors (grazing management, initial disturbance and their interaction) on species richness (S), evenness (accoding to Pielou, 1966) and composition, and also on the ≈ we considered the spring surveys of month 29 (Focht and Borges de Medeiros, 2012) and month 77. For this we used analysis of variance combined with restricted permutation testing (Pillar, 2013) using 10,000 permutations and considering the split-plot design. This randomization (permutation) method is described in Pillar and Orlóci (1996) (see also Anderson, 2001; Manly, 2007) and is implemented in Multiv software, available at http://ecoqua.ecologia.ufrgs.br/ecoqua/MULTIV.html. It generates by data permutation the probabilities for assessing the statistical significance of treatment effects, avoiding in this way the distributional assumptions of the classical F test. The procedure is the same for univariate or multivariate data, since the sum of squares within and between treatments is computed on a distance matrix comparing the relevant experimental units pairwise. For univariate analyses (ANOVA, with species richness and evenness, and E. plana cover) we used Euclidean distances. For multivariate analysis of variance (MANOVA, with species composition, Pillar and Orlóci, 1996) we used chord distances (Legendre and Legendre, 2012). We adopted as test criterion a pseudo-F statistic (Anderson, 2001; Pillar, 2013), which is the ratio of between/within sum of squares, with no consideration of classical ANOVA degrees of freedom. Degrees of freedom are not necessary in permutation testing, for the P-values are generated by permutation and not taken from a theoretical F distribution with known degrees of freedom. The permutations are among observations (ANOVA) or vectors of observations (MANOVA), keeping unchanged the corresponding treatment combination of the experimental unit. However, the procedure in this case, which was a split-plot, block design, required proper restriction of permutations control for the block effects (Pillar and Orlóci, 1996) and to avoid pseudo-replication (Crawley, 2013). Thus, for testing the effects of grazing management, the data of the three subplots in each main plot were permuted altogether among main plots within each block. For testing the effects of initial disturbance and its interaction with grazing management, the permutations were restricted within each main plot (grazing management).
We analyzed the temporal trajectory of species composition for all the treatments by using Principal Coordinates Analysis (PCoA) based on chord distances (Kenkel and Orloci, 1986) using the average species cover values of the treatments in each of the spring surveys, and depicted the trajectories in ordination space. We tested the stability of the ordination axes by bootstrap resampling for 15 axes. These analyses were performed using Multiv software.
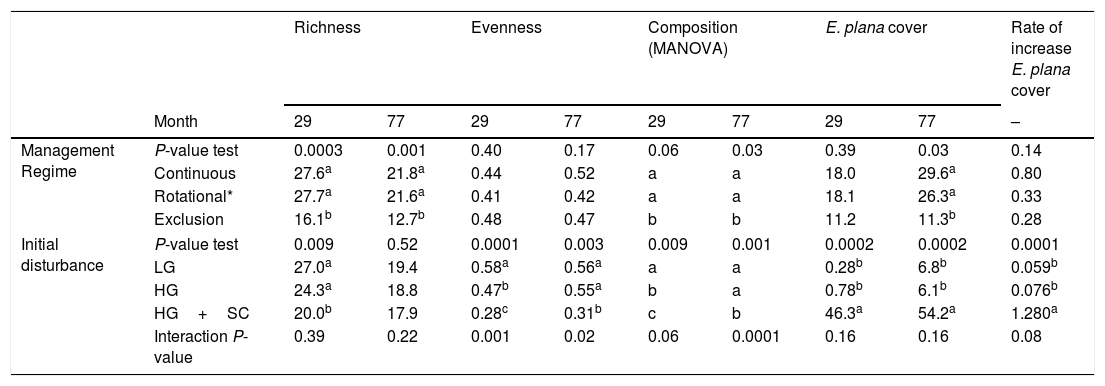
ResultsA total of 140 plant species belonging to 27 families, mostly Poaceae, Asteraceae, Cyperaceae and Apiaceae, was found in all plots and surveys. The mean number of species per survey was 65.9 (± 10). Full data with species composition are provided in electronic Appendix S2. The effect of grazing management on species composition was significant for months 29 and 77, in which grazing exclusion reduced the number of species (<13 species, Table 1). The effect of initial disturbance was highly significant on species richness and on evenness by month 29 (P=0.009 and 0.0001, respectively). By month 77, the effect of initial disturbance was only observed on evenness (P=0.003, Table 1), where the plots that were subjected to heavy grazing plus soil scarification (HG+SC) maintained lower levels of evenness than other initial disturbances. The interaction effect of grazing management and initial disturbance was significant for evenness. For the month 29, the initial disturbance heavy grazing plus soil scarification (HG+SC) combined with the grazing exclusion presented greater evenness than the other grazing managements (Table 2).
Synthesis of community parameters according to the treatments of both evaluated factors (‘management regime’ and ‘initial disturbance’) considering differences in two periods: November 2006 and November 2010 (month 29 and 77, respectively). Mean values of diversity indices (richness and evenness), E. plana cover and its rate of increase are given for each treatment at each period. Values with different letters within each column indicate significant differences (P<0.05) between treatments based on randomization testing for pairwise contrasts. Species composition differences are indicated by different letters within each column (after MANOVA based on randomization testing for pairwise contrasts). Contrasts between treatments were done within each factor. Significant interactions between factors are shown in the last line (P<0.05) and are detailed in Table 2.
| Richness | Evenness | Composition (MANOVA) | E. plana cover | Rate of increase E. plana cover | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Month | 29 | 77 | 29 | 77 | 29 | 77 | 29 | 77 | – | |
| Management Regime | P-value test | 0.0003 | 0.001 | 0.40 | 0.17 | 0.06 | 0.03 | 0.39 | 0.03 | 0.14 |
| Continuous | 27.6a | 21.8a | 0.44 | 0.52 | a | a | 18.0 | 29.6a | 0.80 | |
| Rotational* | 27.7a | 21.6a | 0.41 | 0.42 | a | a | 18.1 | 26.3a | 0.33 | |
| Exclusion | 16.1b | 12.7b | 0.48 | 0.47 | b | b | 11.2 | 11.3b | 0.28 | |
| Initial disturbance | P-value test | 0.009 | 0.52 | 0.0001 | 0.003 | 0.009 | 0.001 | 0.0002 | 0.0002 | 0.0001 |
| LG | 27.0a | 19.4 | 0.58a | 0.56a | a | a | 0.28b | 6.8b | 0.059b | |
| HG | 24.3a | 18.8 | 0.47b | 0.55a | b | a | 0.78b | 6.1b | 0.076b | |
| HG+SC | 20.0b | 17.9 | 0.28c | 0.31b | c | b | 46.3a | 54.2a | 1.280a | |
| Interaction P-value | 0.39 | 0.22 | 0.001 | 0.02 | 0.06 | 0.0001 | 0.16 | 0.16 | 0.08 | |
LG: light grazing; HG: heavy grazing; HG+SC: heavy grazing plus soil scarification; * after month 34 the grazing rotational regime was modified to continuous (see text for details).
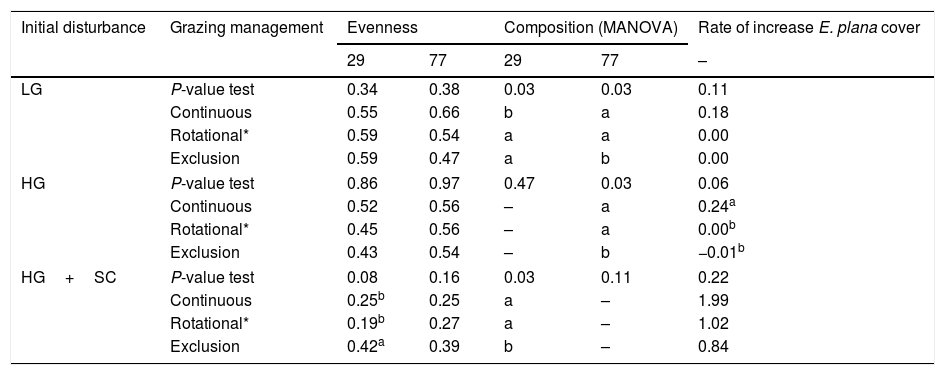
Synthesis of the community parameters that presented interaction between both factors (‘management regime’ and ‘initial disturbance’) shown in Table 1. Values of evenness and rate of increase of E. plana cover correspond to mean values within each combination of treatments of both factors, considering two periods: November 2006 and November 2010 (month 29 and 77 respectively). Values with different letters within a column, within each treatment of the ‘initial disturbance’ factor indicate significant differences (P<0.05) based on randomization testing for pairwise contrasts. Species composition differences are just indicated by different letters within each column.
| Initial disturbance | Grazing management | Evenness | Composition (MANOVA) | Rate of increase E. plana cover | ||
|---|---|---|---|---|---|---|
| 29 | 77 | 29 | 77 | – | ||
| LG | P-value test | 0.34 | 0.38 | 0.03 | 0.03 | 0.11 |
| Continuous | 0.55 | 0.66 | b | a | 0.18 | |
| Rotational* | 0.59 | 0.54 | a | a | 0.00 | |
| Exclusion | 0.59 | 0.47 | a | b | 0.00 | |
| HG | P-value test | 0.86 | 0.97 | 0.47 | 0.03 | 0.06 |
| Continuous | 0.52 | 0.56 | – | a | 0.24a | |
| Rotational* | 0.45 | 0.56 | – | a | 0.00b | |
| Exclusion | 0.43 | 0.54 | – | b | −0.01b | |
| HG+SC | P-value test | 0.08 | 0.16 | 0.03 | 0.11 | 0.22 |
| Continuous | 0.25b | 0.25 | a | – | 1.99 | |
| Rotational* | 0.19b | 0.27 | a | – | 1.02 | |
| Exclusion | 0.42a | 0.39 | b | – | 0.84 | |
LG: light grazing; HG: heavy grazing; HG+SC: heavy grazing plus soil scarification; * after month 34 the grazing rotational regime was modified to continuous (see text for details).
A total of 140 plant species belonging to 27 families, mostly Poaceae, Asteraceae, Cyperaceae and Apiaceae, was found in all plots and surveys. The mean number of species per survey was 65.9 (± 10). Full data with species composition are provided in electronic Appendix S2. The effect of grazing management on species composition was significant for months 29 and 77, in which grazing exclusion reduced the number of species (<13 species, Table 1). The effect of initial disturbance was highly significant on species richness and on evenness by month 29 (P=0.009 and 0.0001, respectively). By month 77, the effect of initial disturbance was only observed on evenness (P=0.003, Table 1), where the plots that were subjected to heavy grazing plus soil scarification (HG+SC) maintained lower levels of evenness than other initial disturbances. The interaction effect of grazing management and initial disturbance was significant for evenness. For the month 29, the initial disturbance heavy grazing plus soil scarification (HG+SC) combined with the grazing exclusion presented greater evenness than the other grazing managements (Table 2).
Synthesis of community parameters according to the treatments of both evaluated factors (‘management regime’ and ‘initial disturbance’) considering differences in two periods: November 2006 and November 2010 (month 29 and 77, respectively). Mean values of diversity indices (richness and evenness), E. plana cover and its rate of increase are given for each treatment at each period. Values with different letters within each column indicate significant differences (P<0.05) between treatments based on randomization testing for pairwise contrasts. Species composition differences are indicated by different letters within each column (after MANOVA based on randomization testing for pairwise contrasts). Contrasts between treatments were done within each factor. Significant interactions between factors are shown in the last line (P<0.05) and are detailed in Table 2.
| Richness | Evenness | Composition (MANOVA) | E. plana cover | Rate of increase E. plana cover | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Month | 29 | 77 | 29 | 77 | 29 | 77 | 29 | 77 | – | |
| Management Regime | P-value test | 0.0003 | 0.001 | 0.40 | 0.17 | 0.06 | 0.03 | 0.39 | 0.03 | 0.14 |
| Continuous | 27.6a | 21.8a | 0.44 | 0.52 | a | a | 18.0 | 29.6a | 0.80 | |
| Rotational* | 27.7a | 21.6a | 0.41 | 0.42 | a | a | 18.1 | 26.3a | 0.33 | |
| Exclusion | 16.1b | 12.7b | 0.48 | 0.47 | b | b | 11.2 | 11.3b | 0.28 | |
| Initial disturbance | P-value test | 0.009 | 0.52 | 0.0001 | 0.003 | 0.009 | 0.001 | 0.0002 | 0.0002 | 0.0001 |
| LG | 27.0a | 19.4 | 0.58a | 0.56a | a | a | 0.28b | 6.8b | 0.059b | |
| HG | 24.3a | 18.8 | 0.47b | 0.55a | b | a | 0.78b | 6.1b | 0.076b | |
| HG+SC | 20.0b | 17.9 | 0.28c | 0.31b | c | b | 46.3a | 54.2a | 1.280a | |
| Interaction P-value | 0.39 | 0.22 | 0.001 | 0.02 | 0.06 | 0.0001 | 0.16 | 0.16 | 0.08 | |
LG: light grazing; HG: heavy grazing; HG+SC: heavy grazing plus soil scarification; * after month 34 the grazing rotational regime was modified to continuous (see text for details).
Synthesis of the community parameters that presented interaction between both factors (‘management regime’ and ‘initial disturbance’) shown in Table 1. Values of evenness and rate of increase of E. plana cover correspond to mean values within each combination of treatments of both factors, considering two periods: November 2006 and November 2010 (month 29 and 77 respectively). Values with different letters within a column, within each treatment of the ‘initial disturbance’ factor indicate significant differences (P<0.05) based on randomization testing for pairwise contrasts. Species composition differences are just indicated by different letters within each column.
| Initial disturbance | Grazing management | Evenness | Composition (MANOVA) | Rate of increase E. plana cover | ||
|---|---|---|---|---|---|---|
| 29 | 77 | 29 | 77 | – | ||
| LG | P-value test | 0.34 | 0.38 | 0.03 | 0.03 | 0.11 |
| Continuous | 0.55 | 0.66 | b | a | 0.18 | |
| Rotational* | 0.59 | 0.54 | a | a | 0.00 | |
| Exclusion | 0.59 | 0.47 | a | b | 0.00 | |
| HG | P-value test | 0.86 | 0.97 | 0.47 | 0.03 | 0.06 |
| Continuous | 0.52 | 0.56 | – | a | 0.24a | |
| Rotational* | 0.45 | 0.56 | – | a | 0.00b | |
| Exclusion | 0.43 | 0.54 | – | b | −0.01b | |
| HG+SC | P-value test | 0.08 | 0.16 | 0.03 | 0.11 | 0.22 |
| Continuous | 0.25b | 0.25 | a | – | 1.99 | |
| Rotational* | 0.19b | 0.27 | a | – | 1.02 | |
| Exclusion | 0.42a | 0.39 | b | – | 0.84 | |
LG: light grazing; HG: heavy grazing; HG+SC: heavy grazing plus soil scarification; * after month 34 the grazing rotational regime was modified to continuous (see text for details).
Regarding species composition, the effect of management regime was marginally significant for the survey conducted at month 29 and significant for month 77 (P=0.06 and P=0.03 respectively, Table 1), when on both communities under grazing exclusion differed in species composition from the other treatments. The effect of initial disturbance on species composition was highly significant at both surveys (29 – P=0.009 and 77 months – 0.001, Table 1). In the survey of month 29 all communities under initial disturbances differentiated between them. As for the survey of the month 77, just the composition of light grazing (LG) and heavy grazing (HG) differed from the heavy grazing plus soil scarification (HG+SC). Interaction between factors also had significant effect on species composition. In the survey of month 29 of the LG treatment, the communities under continuous grazing management differed from the others (P=0.03). There were no significant differences within HG. However, in month 29 to HG+SC, the communities under grazing exclusion differed from other grazing managements (P=0.03, Table 2). The same pattern was also found in month 77, between communities submitted to LG and HG (P=0.03 and P=0.03, respectively).
Main trends in species composition change over time present in Fig. 1. Under grazing exclusion, communities without soil scarification were more stable (shorter trajectories) than those under rotational and continuous grazing, whose trajectories changed towards a higher proportion of E. plana. The first ordination axis (which was stable by the bootstrap test, probabilities P=0.04) depicts increasing E. plana invasion from right to left, which was most evident in the communities subjected to HG+SC, irrespective of grazing management. The second ordination axis indicate, from bottom up, communities freshly modified by initial disturbance, characterized by L. corniculatus, Anagalis arvensis, Lollium multiflorum, Rumex obtusifolius and litter, to grazing exclusion, characterized by Stipa setigera and Paspalum plicatulum. Among treatments submitted to sowing of exotic forage species (L. multiflorum and L. corniculatus and T. repens), in the first survey after the application of the initial disturbances only the plots under HG+SC combined to the two grazing management presented considerable correlation with these species. Correlations dropped in subsequent surveys. More importantly, in the last survey coverage of these species was in average among all treatments only 0.03% for L. multiflorum, and only 0.0016% for both L. corniculatus and T. repens, which reflects their transitory coverage on such grasslands when no recurrent sowing is done.
 of plant communities based on average species composition per treatment recorded in four surveys at 4, 17, 29 and 77 months after the beginning of the experiment. Ordination is based on chord distances between experimental units. The diagrams depict the nine treatments at the four surveys, and the species with the highest correlations with the ordination axes (a) while in the other diagrams the temporal trajectories for each grazing treatment are indicated separately for continuous grazing (b), rotational grazing (c), and grazing exclusion (d). Axis I and II represented 23 and 11% of total variation, respectively. Species shown in (a) are those with a minimum correlation of 0.4 with one of the two axes, i.e.: Paspalum plicatulum (papl – caespitose), Melica brasiliana (mebr – caespitose), Eragrostis plana (erpl – caespitose), Piptochaetium montevidense (pimo – caespitose), Lolium multiflorum (lomu – caespitose), Lotus corniculatus (loco – forb), Anagalis arvensis (anar – forb), Stipa sertigera (stse – caespitose) Sida rhombifolia (sirh -forb), Eragrostis bahiensis (erba – caespitose), Rumex obtusifolius (ruob – forb), Paspalum dilatatum (padi – prostrated).")
Principal Coordinate Analysis (PCA) of plant communities based on average species composition per treatment recorded in four surveys at 4, 17, 29 and 77 months after the beginning of the experiment. Ordination is based on chord distances between experimental units. The diagrams depict the nine treatments at the four surveys, and the species with the highest correlations with the ordination axes (a) while in the other diagrams the temporal trajectories for each grazing treatment are indicated separately for continuous grazing (b), rotational grazing (c), and grazing exclusion (d). Axis I and II represented 23 and 11% of total variation, respectively. Species shown in (a) are those with a minimum correlation of 0.4 with one of the two axes, i.e.: Paspalum plicatulum (papl – caespitose), Melica brasiliana (mebr – caespitose), Eragrostis plana (erpl – caespitose), Piptochaetium montevidense (pimo – caespitose), Lolium multiflorum (lomu – caespitose), Lotus corniculatus (loco – forb), Anagalis arvensis (anar – forb), Stipa sertigera (stse – caespitose) Sida rhombifolia (sirh -forb), Eragrostis bahiensis (erba – caespitose), Rumex obtusifolius (ruob – forb), Paspalum dilatatum (padi – prostrated).
The effect of initial disturbance on invasion was more prominent than management, as hypothesized. Under HG+SC, E. plana cover (54.2%) was namely 8–9 times greater than the other initial disturbances (Table 1). The HG+SC also presented higher rate of increase E. plana cover in the first 34 months. Interaction between factors had marginally significant effect on species composition rate of E. plana cover increase (0.08). The combination of HG and continuous grazing management promoted higher invasion rates compared to other grazing treatments. No differences were detected for the rate of increase of E. plana cover for the interaction between continuous, rotational or exclusion of grazing with initial disturbances of LG or HG+SC (Table 2).
Under grazing exclusion, invasion by E. plana was observed only under HG+SC initial disturbances, where it reached almost 37% of cover in 24 months (Fig. 2). In month 77, cover of E. plana was lowest (±11.3%, P value=0.03) under grazing exclusion compared to other grazing managements (±se 29.6 for continuous and ±23.3% rotational/continuous grazing management, Table 1). The combination of the continuous and rotational/continuous grazing managements and the initial disturbances LG and HG promoted low E. plana coverage as well as low rate of increase E. plana cover during the analyzed period (Table 2 and Fig. 2).
 and three levels of initial disturbance (light grazing, heavy grazing, soil scarification). Propagule pressure was standardized in all treatments. Regression lines and equations are shown for the significant temporal trends under initial disturbance of soil scarification (Continuous: y=1.77x, R2=0.88, P=0.001; Rotational: y=1.58x, R2=0.62, P=0.01; Exclusion: y=1.09, R2=0.46, P=0.01) modelled only during the initial 34 months, before rotational grazing treatment changed to continuous grazing. The bars represent standard deviation of the mean.")
Temporal dynamics of E. plana invasion in communities under three grazing management systems (continuous, rotational and exclusion) and three levels of initial disturbance (light grazing, heavy grazing, soil scarification). Propagule pressure was standardized in all treatments. Regression lines and equations are shown for the significant temporal trends under initial disturbance of soil scarification (Continuous: y=1.77x, R2=0.88, P=0.001; Rotational: y=1.58x, R2=0.62, P=0.01; Exclusion: y=1.09, R2=0.46, P=0.01) modelled only during the initial 34 months, before rotational grazing treatment changed to continuous grazing. The bars represent standard deviation of the mean.
Our results demonstrated the effect of grazing management and disturbances on the temporal dynamics of grassland vegetation facing invasion by E. plana. In plots with similar E. plana propagule pressure, initial disturbance conditions and following grazing management were significant factors that led to differences in the rate of invasion by E. plana, as well as in species composition and diversity.
Community dominance by Poaceae and Asteraceae species is a characteristic of southern Brazilian grasslands (Boldrini, 2009). Despite major changes in the composition of communities due to the invasion of the E. plana, this characteristic remained stable. However, the proportions of species shifted markedly over time and under different combinations of treatments. Species number and evenness decreased in sites where E. plana became abundant, especially in plots that were at the beginning subjected to heavy grazing plus soil scarification. This disturbance was followed by a sharp decline in the performance of perennial grasses (i.e. Paspalum dilatatum and P. plicatulum), which were mostly not able to persist in or recolonize the communities, and by a simultaneous increase of ruderal species, likely available in the soil seed bank, such as R. obtusifolius and A. arvensis (Fig. 1). Among the exotic forage species sown after the application of the initial disturbance, only in plots with heavy grazing plus soil scarification L. multiflorum and L. corniculatus developed expressively in the first surveys. Already in the second survey onwards, there was a change in the communities towards a greater coverage of E. plana and a considerable reduction in the coverage of the sown forage species and of ruderal species.
The heavy grazing plus soil scarification treatment promoted the greatest weakness of the community and, consequently, increased the resource availability. The explanation for this intense change in the community may be the result of a potentiating effect of the interaction between heavy grazing and soil scarification. The heavy grazing technique is also used by livestock producers to establish exotic forage species, which release space at the soil surface (Fitter and Jennings, 1975). As seen, the sown forage species were expressive in the communities only for a short time when compared to E. plana. This short permanence is possibly due to their susceptibility to drought and greater palatability, which may confer less competitive skills in relation to E. plana and some native species. Thus, we think the combination of heavy grazing with soil scarification was predominant over the fact of exotic forage sowing on this treatment. Such initial conditions seem to have affected the whole community structure, persisting over the observation time, and conditioning community trajectory. The soil scarification abruptly breaks community structure and exposes a large proportion of soil, thus providing nutrients and light. Under previous heavy grazing, a greater difficulty to re-establish of pre-existing species is expected because plants may have lower levels of reserves due to the continuous development of leaves that are assiduously consumed by the animals. This combination of treatments probably diminishes the “biological barrier” found in communities under the other initial disturbances (Going et al., 2009), maintaining only the limitations of environmental origin. Thus, opening opportunities for species adapted to local environmental conditions and with attributes linked to the use of resources in the most efficient way for a successful invasion (Grime, 1977).
The intense establishment of E. plana in communities subjected to heavy grazing plus soil scarification may have been exacerbated by the post treatment drought of 2004–2005 (see Focht and Borges de Medeiros, 2012), due to its ability to establish under limited water or nutrients (Scheffer-Basso et al., 2012), and its phenotypic plasticity and conservative strategy of resource use compared to other native species (Coley et al., 1985; Farrior et al., 2013; Favaretto et al., 2015b). Under drought conditions, grazing likely promoted intense chronic herbivory on more palatable species, which favoured plants with defence mechanisms to herbivory (Milchunas and Noy-Meir, 2002). Moreover, this herbivory defence attribute is positively correlated with the ability of plants to tolerate stresses caused by drought (Milchunas et al., 1988). In other words, in periods of drought stress and intense grazing such as those manifested during the first year, E. plana possibly manifests both competitive advantages.
The low cover of E. plana in grazing exclusion is perhaps the result of an accumulation of standing dead biomass (see Focht and Borges de Medeiros, 2012). This accumulation may change physical and biochemical characteristics of the soil surface, constraining the establishment and development of some species, including the invader E. plana. When large tussock species dominate, they tend to competitively exclude species that require light and space close to the soil surface, decreasing the richness of grassland communities (Segre et al., 2016). In our study, the coverage of caespitose species (e.g. P. plicatulum, S. setigera and Mellica brasiliana, Fig. 1) in plots subjected to grazing exclusion increased, while the cover of prostrated (e.g., Paspalum notatum and P. dilatatum) and forbs species (Fig. 1) decreased. Besides its poor adaptation to shady environments like most C4 grasses (Pearcy and Ehleringer, 1984), E plana establishment may also have been, constrained by factors related to high biomass accumulation, low understory temperature, and high soil moisture.
There were only differences for the grazing management in the E. plana rate of increase. No significant changes were detected in the communities as well as for E. plana coverage. Therefore, these plant communities are possibly as much adapted to changes in the frequency and intensity of grazing, as they are dependent on it. Moreover, the inability of the E. plana to invade these communities more intensely is possibly due to the greater use of resources by the set of species already established in these communities. These natural grasslands which coevolved with grazing by large herbivores (Díaz et al., 2007; MacFadden, 2013), presenting species with adaptations to grazing such as protected meristems and high regrowth capacity. Therefore, under grazing, plants of such communities may underutilize some of the existing resources such as soil, light, and nutrients (Moles et al., 2012; Smith and Knapp, 1999; Veen et al., 2008). And these resources provide just a few ‘windows of opportunities’ for E. plana invasion (or even ruderal species) (Davis et al., 2000), which probably limited their recruitment under rotational and continuous treatments.
The soil scarification technique is widely used in the planting of exotic forage species as well as for conversion of grasslands into crop fields or woody plantations. In order to avoid invasion by E. plana, it is utmost important prevent the use of techniques that drastically modify soil conditions such as the soil scarification in grasslands of southern Brazil. And yet, the invasion process can be largely prevented on grasslands by employing either rotational or continuous grazing managements. Removing livestock from grassland is not recommended as it alters drastically the composition and diversity of these ecosystems. Our results are important for devising grazing management strategies of native grasslands that maintain high levels of plant diversity and community resistance to invasion, while providing ecosystem services to society such as livestock production.
The authors thank the Graduate Programs in Animal Science and Ecology of Universidade Federal do Rio Grande do Sul for supporting this research, the Rural Syndicate of Dom Pedrito, Estância Guatambú and EMBRAPA Pecuária Sul for financial and logistical supports. SM and VP have been supported by CNPq (Grants 307719/2012-0 and 307689/2014-0, respectively).
The following are the supplementary data to this article:
Experimental design.
Vegetation data obtained in the experimental plots at four spring surveys (Oct. 2004, Nov. 2005, Nov. 2006 and Nov. 2010).

