


Comparative approaches between native and invasive species germination are useful for understanding invasion success. We studied the role of invasive–native neighboring seeds on germination and early growth of different grasses. We focused on the non-native species Eragrostis plana Ness, the most invasive plant in southern Brazilian grasslands. A germination experiment was performed with three neighboring seed treatments: (i) each species separately, (ii) interspecific seed interactions of native vs. invasive, and (iii) native vs. other native species. The experiment was conducted in a growth chamber; seeds were placed in Petri dishes and grown for 10 days. For each treatment we measured: germination percentage, germination rate index, seedling leaf and root lengths. E. plana germination was remarkably different from native grasses, 73.3% of seeds germinated within 24h. This earlier germination may explain its high invasion potential, as immediate access to limited resources enhances opportunities for establishment and growth. E. plana seeds did not significantly affect germination of other native seeds. However, we demonstrated that some native seeds delayed E. plana germination and decreased its growth. Short delays in germination and growth reduction can result in large differences in final biomass, which may have important implications for preventing E. plana invasion.
Comparisons between native and invasive species performance are useful for understanding what makes an invasive species establishment successful (Daehler, 2003). Plants with certain traits, such as fast and profuse germination, fast growth, and high reproductive ability, are likely to become invasive when introduced to new environments (Baker, 1974). In this context, evaluating germination differences between invasive and native species is a key step to determine if there are intrinsic characteristics that may give invasive species some advantages for colonizing new sites (Schlaepfer et al., 2010). In competitive conditions, invasive species that germinate earlier may have advantages in survival and growth, through space pre-emption and greater access to resources, which could increase the probability of successful establishment.
Invasive species germination rate and emergence might have important consequences for co-occurring native species. Under appropriate conditions, seeds germinate and seedlings compete for resources immediately, modifying water, light and space availability for neighboring seeds. Moreover, seeds may also influence neighboring species by, for example, releasing chemical inhibitors into their surroundings (Friedman and Waller, 1983; Tang and Zhang, 1986; Wardle et al., 1991). This effect has been documented more for invasive plants, where invasive species seeds produce chemical compounds that can affect other plants performance (Wardle et al., 1991; Laterra and Bazzalo, 1999). However, little is known about the influence of invasive species seeds on neighboring native seeds, as a potential strategy for successful colonization and establishment. Evaluating the effect of an invasive species seed on neighboring seeds could have important implications for understanding how invasive species impact native species germination and early emergence. In addition, native seeds could have some influence on co-occurring seeds of an invasive species, e.g. decreasing its germination and early growth rates, which may have important implications for restoration strategies.
Eragrostis plana Ness (Poaceae; “capim Annoni”), a perennial grass introduced from South Africa, is the most invasive plant in southern Brazilian natural grasslands (Medeiros and Focht, 2007; Guido and Guadagnin, 2015; Guido et al., 2016). It is also considered invasive in other regions of South America, including Uruguay and the Mesopotamia region in northeastern Argentina (Barbosa et al., 2013). In this study, we performed a short-term germination experiment to explore some mechanisms that may explain the success of E. plana invasion in southern Brazil. Specifically, we (i) compared the germination rates between E. plana and eight common native grasses, (ii) evaluated the effect of E. plana seeds on native species germination and early growth (i.e. leaves and root length), and (iii) investigated the influence of native neighboring seeds on E. plana success.
MethodsSeed collectionWe collected mature seeds of nine C4 perennial grasses, commonly distributed in natural grasslands in the state of Rio Grande do Sul, southern Brazil (Appendix S1). Eight species were native: Aristida laevis (Ness) Kunth, Axonopus affinis Chase, Eragrostis lugens Nees, Eragrostis neesii Trin., Paspalum notatum Flüggé, Setaria parviflora (Poir.) Kerguélen, Steinchisma hians (Elliott) Nash and Sporobolus indicus (L.) R. Br.; and one invasive: E. plana. Seeds were dried naturally, separated manually and checked for healthy appearance (i.e. only those without evidence of herbivory and fungi were included). For each species, we randomly selected seeds to be used in the experiment.
Experimental designWe conducted a germination experiment in Petri dishes with three neighboring seed treatments (n=4): (i) each species separately (intraspecific seed interaction), and, for each native grass species, interspecific seed interactions of (ii) native vs. E. plana and (iii) native vs. other native species, with a constant density of 30 seeds per Petri dish (see Appendix S2). We selected S. indicus as the other native species in treatment (iii), as this grass presents a growth form similar to E. plana. This treatment is crucial for controlling for the effect of the neighboring seed species identity, as it can disentangle the impacts of native and invasive species. For both interspecific seed interaction treatments (ii) and (iii), 15 seeds of each species were interspersed and uniformly distributed in the Petri dish.
The experiment was conducted in a sterilized system with two layers of filter paper in a Petri dish moistened with 5mL of distilled water. Petri dishes were sealed and placed randomly in a growth chamber with a constant temperature of 25°C and a photoperiod of 12h for 10 days. Each experimental unit consisted of 30 seeds forming a grid of five rows by six columns. Germinated seeds in each Petri dish were counted daily to calculate germination percentage, beginning on the first day the experiment was installed.
On the last day of the experiment, we measured the average leaf length (i.e. cotyledon) and root length of seedlings in each Petri dish. For that, we randomly sampled ten seedlings for the individual species treatment (intraspecific treatment) and five seedlings of each species in mixtures (interspecific treatments) per Petri dish.
We also calculated the germination rate index (GRI) for the 10 days interval as:
where i is the germination count day, varying from day 2 to day 10, Gi is the percentage of seeds germinated by day i, and Gi−1 is the percentage of seeds germinated after the previous count day (Maguire, 1962).Data analyses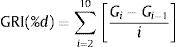
We used ANOVAs to compare: (i) the germination percentage between species; (ii) the neighboring seed treatments; and (iii) the effects of native neighboring seeds on E. plana germination and growth. ANOVAs with permutation testing (Manly, 2007) was performed with MULTIV software (available at http://ecoqua.ecologia.ufrgs.br). For all permutation tests we used 10,000 permutations.
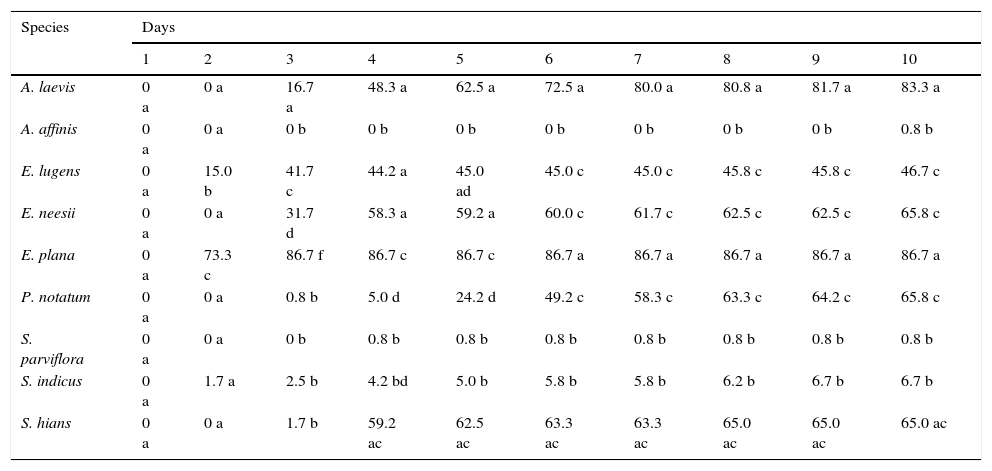
ResultsWe observed significant differences between species germination throughout the experiment (Table 1). Germination of E. plana was remarkably high, reaching 73.3% on the second day of the experiment (24h after seeding). This value increased to 86.7% on the third day, and then remained constant until the end of the experiment. A. laevis (83.3%) was the native grass that achieved similar total germination values to E. plana. E. lugens (46.7%), E. neesii (65.8%) and P. notatum (65.8%) had intermediate values of total germination. A. affinis (0.8%), S. parviflora (0.8%) and S. indicus (6.7%) were the species with lowest germination values by the end of the experiment.
Germination percentage (%) for each species during the 10 days of the experiment (only results of the intraspecific seed interaction treatment are shown). Seed density was constant (30 seeds per Petri dish; n=4). Same letters indicate non-significant differences (P>0.05) between species within each day of the experiment.
| Species | Days | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |
| A. laevis | 0 a | 0 a | 16.7 a | 48.3 a | 62.5 a | 72.5 a | 80.0 a | 80.8 a | 81.7 a | 83.3 a |
| A. affinis | 0 a | 0 a | 0 b | 0 b | 0 b | 0 b | 0 b | 0 b | 0 b | 0.8 b |
| E. lugens | 0 a | 15.0 b | 41.7 c | 44.2 a | 45.0 ad | 45.0 c | 45.0 c | 45.8 c | 45.8 c | 46.7 c |
| E. neesii | 0 a | 0 a | 31.7 d | 58.3 a | 59.2 a | 60.0 c | 61.7 c | 62.5 c | 62.5 c | 65.8 c |
| E. plana | 0 a | 73.3 c | 86.7 f | 86.7 c | 86.7 c | 86.7 a | 86.7 a | 86.7 a | 86.7 a | 86.7 a |
| P. notatum | 0 a | 0 a | 0.8 b | 5.0 d | 24.2 d | 49.2 c | 58.3 c | 63.3 c | 64.2 c | 65.8 c |
| S. parviflora | 0 a | 0 a | 0 b | 0.8 b | 0.8 b | 0.8 b | 0.8 b | 0.8 b | 0.8 b | 0.8 b |
| S. indicus | 0 a | 1.7 a | 2.5 b | 4.2 bd | 5.0 b | 5.8 b | 5.8 b | 6.2 b | 6.7 b | 6.7 b |
| S. hians | 0 a | 0 a | 1.7 b | 59.2 ac | 62.5 ac | 63.3 ac | 63.3 ac | 65.0 ac | 65.0 ac | 65.0 ac |
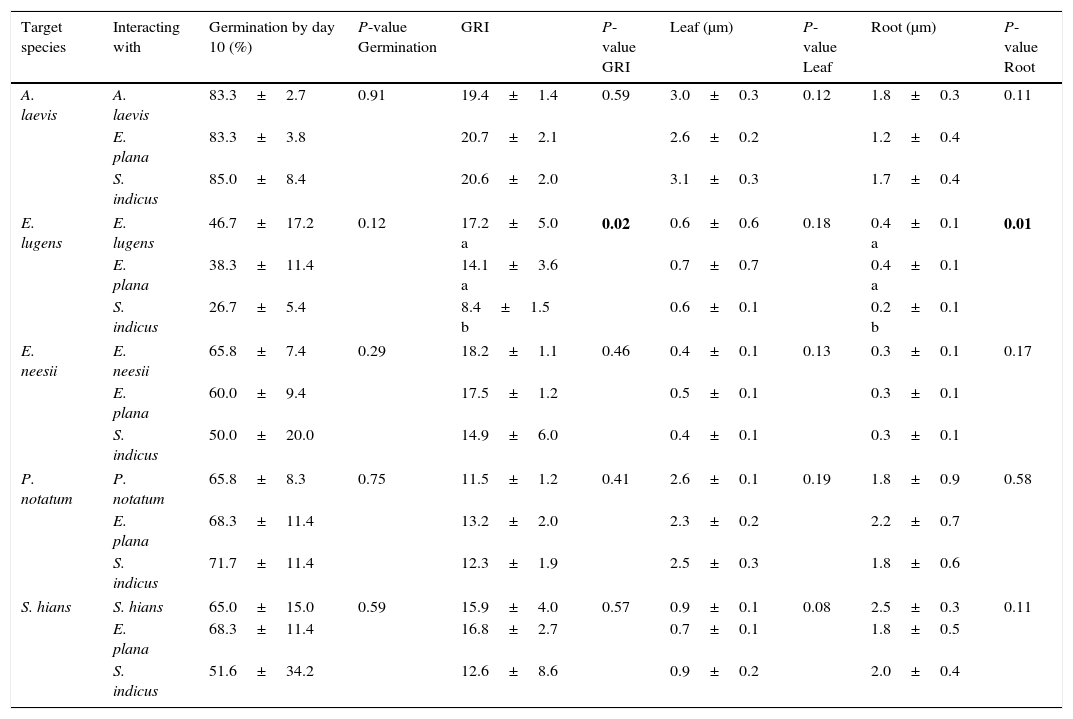
We did not find a significant effect of neighboring seed treatments for native grasses germination or early leaf length (Table 2). This means that the species identity of neighboring seeds did not influence total germination or early growth of leaves of each native grass. However, we found that GRI and root length for E. lugens was reduced in the presence of S. indicus seeds. A. affinis, S. parviflora and S. indicus were not considered in this analysis as target species, due to very low values of total germination.
Germination percentage (%), germination rate index (GRI), leaf and root length (μm) of target species subject to three neighboring seed treatments (intraspecific, and interspecific native vs. E. plana and native vs. another native) after 10 days of the experiment for each target species. The density of seeds was constant among treatments (30 seeds; number of Petri dishes per treatment: n=4). P-values in bold show significant difference (P≤0.05) between treatments for each target species. Same letters indicate non-significant differences (P>0.05) between species.
| Target species | Interacting with | Germination by day 10 (%) | P-value Germination | GRI | P-value GRI | Leaf (μm) | P-value Leaf | Root (μm) | P-value Root |
|---|---|---|---|---|---|---|---|---|---|
| A. laevis | A. laevis | 83.3±2.7 | 0.91 | 19.4±1.4 | 0.59 | 3.0±0.3 | 0.12 | 1.8±0.3 | 0.11 |
| E. plana | 83.3±3.8 | 20.7±2.1 | 2.6±0.2 | 1.2±0.4 | |||||
| S. indicus | 85.0±8.4 | 20.6±2.0 | 3.1±0.3 | 1.7±0.4 | |||||
| E. lugens | E. lugens | 46.7±17.2 | 0.12 | 17.2±5.0 a | 0.02 | 0.6±0.6 | 0.18 | 0.4±0.1 a | 0.01 |
| E. plana | 38.3±11.4 | 14.1±3.6 a | 0.7±0.7 | 0.4±0.1 a | |||||
| S. indicus | 26.7±5.4 | 8.4±1.5 b | 0.6±0.1 | 0.2±0.1 b | |||||
| E. neesii | E. neesii | 65.8±7.4 | 0.29 | 18.2±1.1 | 0.46 | 0.4±0.1 | 0.13 | 0.3±0.1 | 0.17 |
| E. plana | 60.0±9.4 | 17.5±1.2 | 0.5±0.1 | 0.3±0.1 | |||||
| S. indicus | 50.0±20.0 | 14.9±6.0 | 0.4±0.1 | 0.3±0.1 | |||||
| P. notatum | P. notatum | 65.8±8.3 | 0.75 | 11.5±1.2 | 0.41 | 2.6±0.1 | 0.19 | 1.8±0.9 | 0.58 |
| E. plana | 68.3±11.4 | 13.2±2.0 | 2.3±0.2 | 2.2±0.7 | |||||
| S. indicus | 71.7±11.4 | 12.3±1.9 | 2.5±0.3 | 1.8±0.6 | |||||
| S. hians | S. hians | 65.0±15.0 | 0.59 | 15.9±4.0 | 0.57 | 0.9±0.1 | 0.08 | 2.5±0.3 | 0.11 |
| E. plana | 68.3±11.4 | 16.8±2.7 | 0.7±0.1 | 1.8±0.5 | |||||
| S. indicus | 51.6±34.2 | 12.6±8.6 | 0.9±0.2 | 2.0±0.4 | |||||
Remarkably, we found that some species delayed the germination of E. plana (Fig. 1). On the second day of the experiment (24h after seeding), whether separately (intraspecific interaction), or in combination with A. laevis or P. notatum, germination of E. plana was in average 79.4%, contrasting with a germination of 12.7% when it was together with E. lugens, S. parviflora, S. hians or E. neesii (Fig. 1). However, total germination at the end of the experiment did not differ between treatments (Figs. 1 and 2a). In addition, the native grasses A. affinis, E. lugens, S. parviflora, S. hians and E. neesii significantly decreased the GRI of the invasive species (Fig. 2b). Moreover, the presence of A. affinis decreased the length of E. plana leaves (Fig. 2c), and E. lugens decreased its root length (Fig. 2d).
 of E. plana over time for each neighboring seed species (E. plana separately and with eight native species) during 10 days of the experiment. The density of seeds was constant in the Petri dishes (30 seeds; n=4).")
, germination rate index (GRI; b), leaf (c) and root length (d) of E. plana for each interaction treatment (E. plana separately and with eight native species) for the 10 days of the experiment. The density of seeds was constant in the Petri dishes (30 seeds; n=4). Same letters indicate non-significant difference (P>0.05) between treatments.")
Germination percentage (a), germination rate index (GRI; b), leaf (c) and root length (d) of E. plana for each interaction treatment (E. plana separately and with eight native species) for the 10 days of the experiment. The density of seeds was constant in the Petri dishes (30 seeds; n=4). Same letters indicate non-significant difference (P>0.05) between treatments.
By using a simple and short-term germination experiment, we demonstrated that E. plana germination is remarkable fast when compared with co-occurring native grasses. This fast germination rate may be an important contributing factor explaining its high invasion potential in southern Brazilian natural grasslands. However, we did not find significant effects of E. plana seeds on the germination and early growth of native grasses. Remarkably, we demonstrated that some neighboring native seeds have the potential to delay E. plana germination and to decrease the length of its leaves and roots, which could have important implications for preventing E. plana colonization and for restoring invaded areas.
Germination percentage and timing are key life-history traits that play an important role in biological invasions (Baker, 1974; Zhang et al., 2011). Our results indicated that in controlled conditions (i.e. constant 25°C temperature and not limited resources), E. plana reaches 73.3% of germination within 24h. This result is important, as early germination might increase plant fitness (Verdú and Traveset, 2005; Schlaepfer et al., 2010), through space pre-emption and immediate access to limited resources, enhancing opportunities for invasion success. However, the proportion of germinated seeds of E. plana (86.7%) was not significantly different from A. laevis (83.3%) by the end of the experiment, which also demonstrated that some native grasses have similar germination percentages as the invasive species.
Although E. plana invasion is currently one of the major ecological and economical threats in southern Brazilian natural grasslands (Medeiros and Focht, 2007; Guido and Guadagnin, 2015; Guido et al., 2016), information about its impact is still scarce. Guido and Pillar (2017) demonstrated its impact on grassland communities by reducing resident species richness and total cover. Other authors have suggested an allelopathic effect of E. plana's leaf and root extracts on native vegetation by affecting the germination rates of other species (Ferreira et al., 2008; Favaretto et al., 2011). However, this observation has not been verified in field conditions. In our study, we did not find any effects of E. plana seeds on germination and early growth of eight native grasses, at least in controlled laboratory conditions. Thus, if E. plana has an inhibitory effect on native vegetation, this may occur at later stages of the invasion process.
Several studies have reported chemical inhibitors in seeds that are often released into their nearby surroundings, affecting germination and/or early growth of potential competitors (Wardle et al., 1991; Laterra and Bazzalo, 1999; Zhang et al., 2011). In our experiment, we demonstrated that some native seeds have the potential to briefly delay E. plana germination, and also to decrease leaf and root lengths. These native grasses were E. lugens, S. parviflora, S. hians and E. neesii, common plants in southern Brazilian grasslands which frequently co-occur with E. plana. These findings are novel and important, as brief delays in germination and early growth can result in large differences in final biomass and reproduction success (Baker, 1974), which may have important implications for invasive species colonization.
As a conclusion, we demonstrated that E. plana seeds have a high (73.3%) germinated rate within 24h and, when grown under optimal conditions for 10 days, the percentage of germinated seeds reached 86.7%. These results reflect an important strategy that enable E. plana to be a successful invasive species in southern Brazilian natural grasslands. We did not find an effect of E. plana seeds on native species germination and growth, suggesting that if there is an inhibitory effect, it may occur during later stages of the invasion process. However, seed germination and emergence may also be strongly influenced by other factors that were not investigated in this experiment, such as maternal effects (Roach and Wulff, 1987), resource limitation and seed density (e.g. Laterra and Bazzalo, 1999). E. plana produces a large number of seeds each reproductive season, so its density in the seed bank may be higher than some native species, which is relevant information to consider in future research. We also demonstrated that some native grasses not only have the potential to delay E. plana germination, but also to decrease its early growth. These results may reveal potential options for restoring invaded areas by reintroducing native seeds to the seed bank, as delays in germination and growth can have important implications for preventing E. plana colonization.
FundingThis work received support from the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, Brazil) provided to A. Guido (grant 402943/2015-5) and V. Pillar (grant 307689/2014-0); and from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES, Brazil).
We thank Eliane da Silva for laboratory material and helpful suggestions during the experiment; and Vasiliki Balogianni and Carolina Schnitzler for helping on data collection.
The following are the supplementary data to this article:

