






The crescent urbanization across the globe has widespread ecological consequences, affecting species distribution, demography and behaviour. In this study we aimed to identify, quantify, map and predict the occurrences of medium- to large-bodied (≥1 kg) mammal carnivorans across urban and peri-urban landscapes throughout Brazil. We use a news-based search to compose a dataset of occurrences across the Brazilian cities. Additionally, we gather various socio-environmental and geophysical variables from these urban areas to identify the drivers of carnivore presence across urban environments. Employing Kernel interpolation, we generate heatmaps to highlight and pinpoint regions in Brazil with the highest concentration of carnivorans species on urban areas. We also employed a Generalized Additive Models (GAM) approach to assess the predictive power of social-environmental and geophysical variables on the occurrence of carnivorans in urban landscapes of Brazil. Our findings pinpoint to an increase of potential human-wildlife conflicts likely given that the urban expansion over natural habitats had a rampant increase in the last decades. Finally, we highlight that our results can serve as a basis to improve urban planning and offer important insights into the intricate interplay of social and geophysical variables that influence the occurrences of carnivorans within urban environments in Brazil.
The widespread phenomenon of global urbanization is experiencing a rapid increase, leading to extensive ecological consequences that surpass the boundaries of urban areas. As pointed out in the report of the United Nations (UN DESA, 2022), it is projected that urban areas will accommodate 60% of the global human population by 2030. Moreover, these projections indicate that approximately 89.3% of Brazilian — the largest tropical country — population will reside in large- (i.e., from 5 to 10 million people) or medium-sized cities (i.e., from 1 to 5 million people), most of them located in coastal region. Consequently, biodiversity is undergoing shifts in response to urban landscape expansions. Several studies indicate that urbanization has a significant ecological impact, primarily resulting in the loss of natural habitats and, subsequently, biodiversity loss or assembly rearrangements (Mckinney, 2002; Magle et al., 2021). Urbanization tends to favour a limited number of species that thrive in urban areas (Mckinney, 2002), classified as urban adapters and exploiters, while other species are classified as urban avoiders (McKinney, 2002).
Terrestrial carnivore mammals (Order Carnivora) are particularly vulnerable to urbanization due to their high spatial and energetic requirements (i.e., home ranges and trophic level), low reproductive rates, and human-wildlife conflicts, for example, persecutions or retaliatory hunting (Barker et al., 2023). However, despite the numerous barriers imposed by the urban environment across its highly modified landscapes, such as decreases in vegetation cover and natural resources, habitat fragmentation and degradation, extensive road networks, and unsuitable climate conditions (Barker et al., 2023), it is intriguing to observe the establishment of several carnivorans in cities worldwide. For example, we highlight the Red foxes (Vulpes vulpes) in Great Britain (Scott et al., 2014), Raccoons (Procyon lotor) — arguably one of the most common carnivores in North American cities (Hadidian et al., 2010), Japan and Germany where it was introduced (see Bateman and Fleming, 2012) —, and Coyotes (Canis latrans) across cities Southwestern North America (Gehrt et al., 2010).
Some carnivorans species exhibit lower reliance on human-provided food sources and instead rely on vast expanses of native vegetation and natural resources (e.g., Jaguar (Panthera onca), Puma (Puma concolor), and Maned Wolf (Chrysocyon brachyurus) (De Angelo et al., 2011; Regolin et al., 2017). However, these large-bodied species are being frequently observed in suburban regions, where relative low human density permits a concentration of larger parcels of natural habitats when compared to urban city cores (Bateman and Fleming, 2012). Studies conducted at several scales of habitat use by carnivorans have consistently demonstrated that the majority of carnivorans tend to avoid intensively anthropized areas (Bateman and Fleming, 2012; Bruskotter et al., 2017; Magle et al., 2021). However, due to the rapid expansion of urban environments, there is an increasing frequency of mammalian carnivores utilizing these modified areas, especially in countries with emergent economies, such as Brazil.
Yet, the large-scale effects of urbanization, land-use changes, and habitat loss and fragmentation on the distribution, occurrence, and dispersal abilities of mammalian carnivorans throughout Brazil remain poorly understood. Crowdsourcing platforms —gathering information from a broad online community — have become powerful tools for capturing extensive ecological data at a relatively low-cost (Grade et al., 2022). This approach is particularly useful to investigate several socio-ecological patterns in cities (Grade et al., 2022). Across a spatial gradient of heterogeneity and intricate land ownership patterns — highly prominent in urban environments — crowdsourcing provides valuable information. However, studies suggest that factors like socio-economic status and segregation, whether driven by poverty or racial disparities, may influence the acquisition of biodiversity data (Ellis-Soto et al., 2023). This trend may also extend to the reporting of occurrences of carnivorans mammals in urban areas, particularly in smaller municipalities. This is because only 14.4% of Brazilian municipalities, where 52.35% of the population resides, have a local TV station (http://sistemas.anatel.gov.br). Yet, to our knowledge, no research to date has employed local news outlets (e.g., crowdsourced platforms) to obtain information on occurrences of carnivorous mammals in urban areas across Brazil.
In this study, we aimed to identify, quantify, map, and predict — according to socio-environmental and geophysical variables — the occurrences of medium- to large-bodied (≥1 kg) carnivorans (Families Felidae and Canidae) across urban and peri-urban landscapes throughout Brazil. We hypothesised that occurrences of medium- to large-bodied carnivorans in urban areas across Brazil are primarily explained by environmental variables (i.e., defaunation index, remaining native habitat cover, proximity with Protected Areas, biomass above ground, and vegetation height) given that these variables represent the ecological components — therefore the major requirements — of species (McKinney, 2002; De Angelo et al., 2011; Bateman and Fleming, 2012; Regolin et al., 2017). Further, the occurrences of carnivorans are secondarily explained by social and geophysical variables (i.e., human development index, human population density, poverty index, road network, elevation profile, and municipality size (km²)) (Frank et al., 2019). Furthermore, we expect that the response of carnivorans — in terms of species occurrence — to socio-environmental and geophysical variables differs among the Brazilian biomes, reflecting, therefore, the levels of urbanization across different regions of Brazil.
Material and methodsData compilationFrom early December 2022 to late July 2023, we used the Google Search engine (www.google.com.br) to search for any news outlets noticing the presence of medium- to large-bodied mammalian carnivorans across Brazil. To do so, we used a set of keywords in Portuguese as follows: “Onça” (i.e., Jaguar or Puma); “Lobo” (i.e., wolf as representing canids); “Puma” (i.e., Cougar, Mountain Lion); “Cão-selvagem” (wild dog to canids); “Gato-selvagem” (small-bodied wild cats, such as Leopardus sp. and Herpailurus yagouaroundi); “Jaguatirica” (i.e., Ocelot) and its regional folk-taxonomy variations, such as “Graxaim” for canids. We then compiled and validated the information about the species and the respective municipality where the record, considering only records from 2010 to 2022 (i.e., storage time on websites of news outlets). Moreover, for each record, we compiled additional information regarding the posting date, sex and age of the species when available, and additional information on the destination of the specimen. Based on this search, we compiled a dataset composed of 605 independent occurrences of 10 species of felids and canids within 368 urban localities across all Brazilian biomes (except Pampa) (Fig. 1a). For details about both common and scientific names of species see Supplementary table S1.
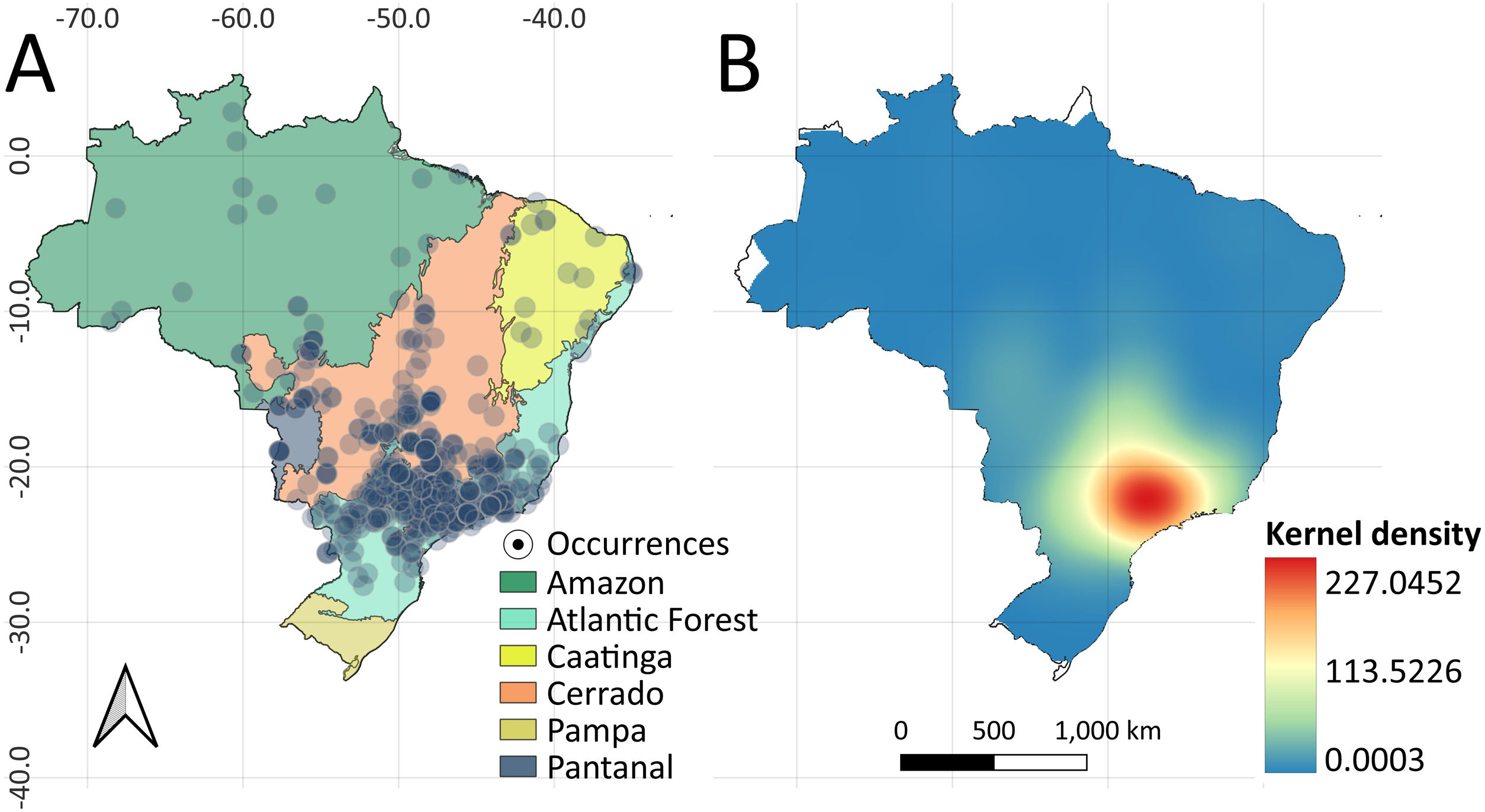
(A) Occurrences of mammalian carnivorans (Felidae and Canidae) across urban areas throughout Brazilian biomes between 2010 and 2022. (B) Kernel interpolation (heatmap) showing the spatial hotspots (highlighted in red) and therefore indicating places with high rates of occurrences of medium- to large-bodied carnivorans in Brazilian urban areas.
Once we compiled the municipalities where each carnivorans records occurred, based on its political limits sourced from the Brazilian Institute of Geography and Statistics (IBGE: https://cidades.ibge.gov.br) and expressed by latlong (SIRGAS 2000) polygons, we obtained multiple social, environmental, and geophysical variables. These variables at municipal-scale, were represented by social aspects: (1) human development index (HDI; 2010); (2) human population density (HPD; 2021); (3) poverty index (2010; representing the proportion of individuals with per capita household monthly income ≤ US$ 27.41) all based on IBGE – Instituto Brasileiro de Geografia e Estatística, 2021 and Brazilian Human Development Atlas (PNUD, 2013). The environmental variables were composed of: (1) mammal defaunation index (municipal-scale mean; sourced in Bogoni et al., 2020); (2) rate of remaining native habitat (all native vegetation classes) cover in 2019 (sourced from MapBiomas 2022); (3) the linear proximity of any protected area (PAs; in km sourced from UNEP-WCMC and IUCN (2022) in relation to the municipality centroid; (4) nocturnal lights hotspots (n; 2021; sourced from NASA Earth Data: https://www.earthdata.nasa.gov); (5) biomass above ground (M.g/ha); and (6) vegetation height (m), both derived from GEDI-Landsat data (i.e., fusion with Landsat maps of vegetation change, available in GLAD Landsat ARD) to provide high spatial resolution estimates of vegetation height and change in aboveground carbon stocks at annual time-scales) (Potapov et al., 2020). In geophysical terms, we extracted: (1) the extension of state and federal roads (km; dated from 2021 and sourced from Departamento Nacional de Infraestrutura de Transportes: https://www.gov.br/dnit/pt-br) over municipality extensions; (2) elevation (meters above sea level [masl]) derived from ASTER-GDEM-NASA); and (3) the municipality size (km²) derived from the Brazilian Institute of Geography and Statistics (IBGE – Instituto Brasileiro de Geografia e Estatística, 2021). Data extractions were performed using R version 4.3.0 (https://www.r-project.org/) and Google Earth’s Engine platform (code.earthengine.google.com). Additionally, for each municipality, the percentage of the major biome to which its territorial area belongs was obtained by overlapping biome vs. municipality polygons.
Social, environmental, and geophysical variables were not autocorrelated, given that the VIF was typically ≤3, except for biomass above ground and vegetation whose VIF was 10, indicating a possible autocorrelation with other variables. Nonetheless, the likelihood ratio test indicated that models without the variables biomass above ground and vegetation height were not significantly different from the overall model, suggesting that these variables may not have a statistically significant impact on the model responses (p < 0.001). The evaluation of the basis dimension for each smoothing term revealed that the terms were adequately adjusted, with k-index values close to 1 and p-values > 0.05.
Data analysisFirst, we used a Kernel interpolation approach (Wand and Jones, 1995) to create heatmaps aiming to depict and identify regions in Brazil with the highest concentration of occurrences of all carnivorans over urban areas. We used the same approach considering each species of canids and felids. Species with less than 10 records were grouped to create a single heatmap. The Kernel approach permits to estimate the number of events per unit area in each cell of a regular grid that covers the study area (Wand and Jones, 1995). To create the heatmaps, we utilized the Kernel Density Estimation tool in QGIS 3.28.3 (QGIS Development Team, 2022) with a radius of 0.5º (∼55-km) around the points and a resolution of 0.1 units per pixel on the x and y axes (Wand and Jones, 1995).
We employed a Generalized Additive Models (GAM) approach to assess the predictive power of social, environmental, and geophysical variables on the occurrence of carnivorans in urban landscapes throughout Brazil. Additionally, we perform GAM models for each species with sufficient occurrence points (i.e., >10) to investigate how predictive variables influences each species. The GAM models were performed by applying smoothed functions (s) to represent the flexible relationship between predictor variables and the response variable (Wood, 2017). GAM models provide a powerful tool for exploring complex relationships, as they avoid rigid parametric assumptions on how predictors influence the outcome, therefore, this statistical approach allows us to capture nuanced patterns in the data without imposing specific functional forms (Wood, 2017). The GAM was specified as follows: response variable (i.e., number of events in any urban area) ∼ s(human population density) + s(human development index) + s(poverty) + s(extension of roads ) + s(elevation) + s(municipality size) + s(defaunation index) + s(rate of remaining native habitat ) + s(proximity of protected area) + s(nocturnal lights hotspots) + s(biomass above ground) + s(vegetation height) + s(Biome, bs = ‘re’). We included a “random effect” based smooth (“bs”) for the biome variable, to account for variations in the number of events across different biomes and regions of Brazil. This allowed for effective control of spatial heterogeneity in the dataset, leading to more accurate and reliable model results. Smoothing parameters were estimated using restricted maximum likelihood. To handle variations in numerical magnitudes and asymmetry of predictor variables, we applied natural log-transformations (i.e., log x + 1) followed by standardization using the “decostand” function from the R package vegan (Oksanen et al., 2022). This preprocessing ensured comparability and improved the models accuracy and interpretability (see Wood, 2017). We tested predictor variables for multicollinearity using an a priori variance inflation factor test (VIF, Dobson 1990) and considered values ≤10 as acceptable before constructing the models. GAMs were performed under a Poisson distribution with a ‘log’ link function under the variance (Wood, 2017). The model was performed using a smoothing factor equal to the number of biomes minus one (i.e., k = 5; Wood, 2017). We performed GAMs in version 4.3.0 (https://www.r-project.org/) based on the mgcv (Wood, 2011) and HH (Heiberger, 2018) R-packages (see Supplementary Material for GAM R-code).
ResultsWe identified 605 occurrences of medium- to large-bodied mammalian carnivorans belonging to 10 species of felids and canids, which were observed across five over six Brazilian biomes, belonging to 368 urban areas (Fig. 1a; see Appendix Tables S2 and S5). The records among the biomes were staggered, with the largest percentage in the Atlantic Forest and the lowest in the Pantanal, while the Pampas had no records (Atlantic Forest [N = 323; frequency % [F] = 53.39]; Cerrado [N = 234; F = 38.68]; Amazon [N = 25; F = 4.13]; Caatinga [N = 12; F = 1.98]; Pantanal [N = 11; F = 1.82]) (see Fig. S1, Table S2 for details). The kernel density estimator indicated an area of 586,265 km² along the southern portion of the Atlantic Forest and Cerrado, comprising the Southeast region of Brazil, with the highest concentration of carnivorans in urban areas, forming an impressive spatial hotspot of occurrences (Fig.1B).
The felids species with the highest occurrence rates were Puma concolor (N = 256; frequency [F] = 42.31), Leopardus pardalis (N = 60; F = 9.92), and Panthera onca (N = 35; F = 5.79), followed by small-bodied felid fauna such as Herpailurus yagouaroundi (N = 4; F = 0.66), Leopardus wiedii (N = 2; F = 0.33%), and L. guttulus/L. tigrinus (N = 1; F = 0.17) (Fig. 2). Among the canids, the species most frequently recorded were Chrysocyon brachyurus (N = 195; F = 32.23), Cerdocyon thous (N = 48; F = 7.93), and Lycalopex sp. with (N = 2; F = 0.33) (Fig. 3). When considering the species individually, there are divergences between the occurrence heatmaps and the overall pattern found when considering all records (Fig. 2 and Fig. 3; Table S1 and S2). The main difference is for P. onca, whose occurrences concentrate in Pantanal-Cerrado-Amazon contact zones (Fig. 2).
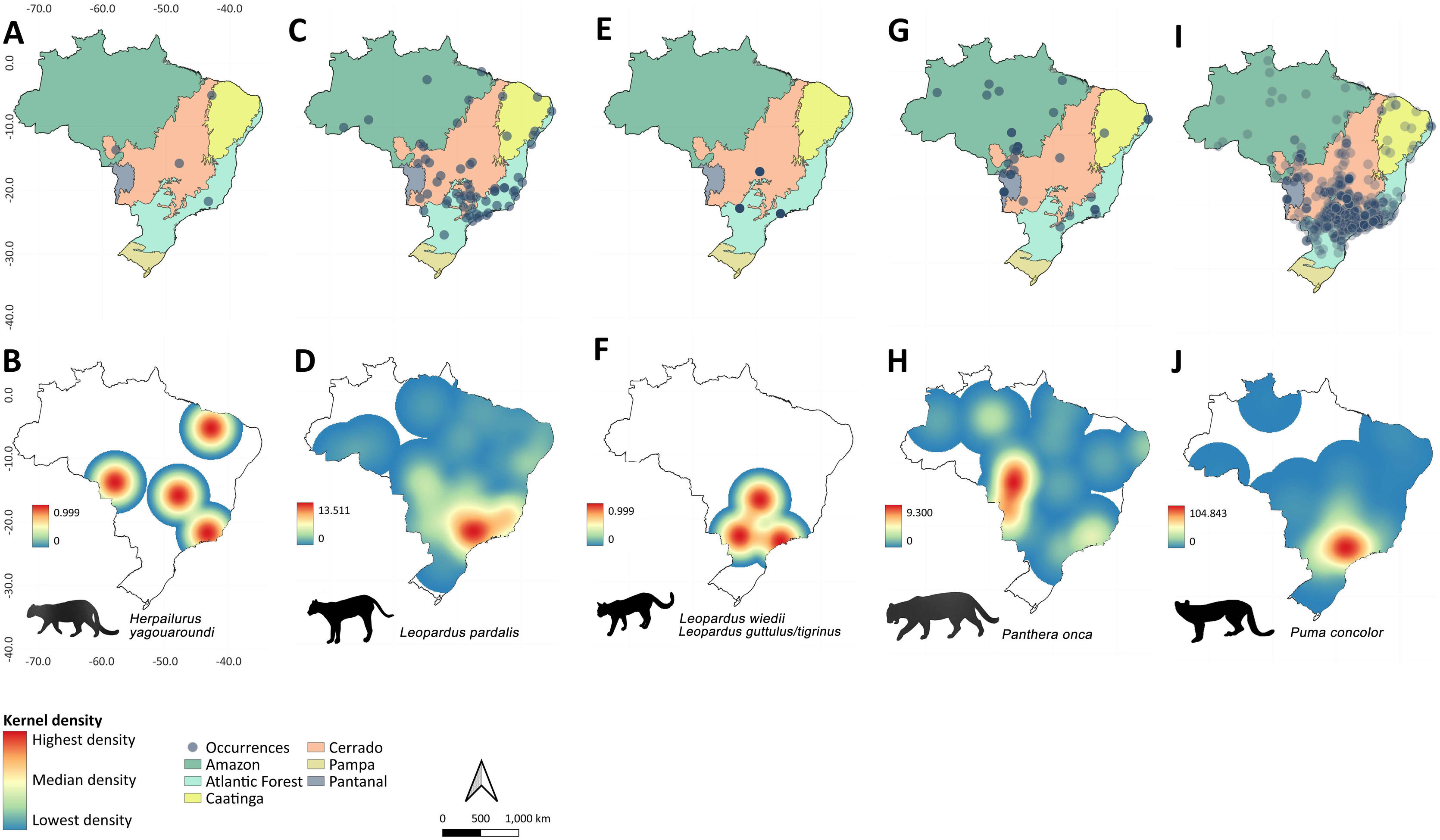
Occurrences of felids in urban areas throughout Brazil between 2010 and 2022, and the respective heatmaps based on Kernel approach showing spatial hotspots (highlighted in red) with high rates of occurrences. Felids species are: Herpailurus yagouaroundi (A-B), Leopardus pardalis (C-D), Leopardus wiedii and Leopardus guttulus/tigrinus (E-F), Panthera onca (G-H) and Puma concolor (I-J).
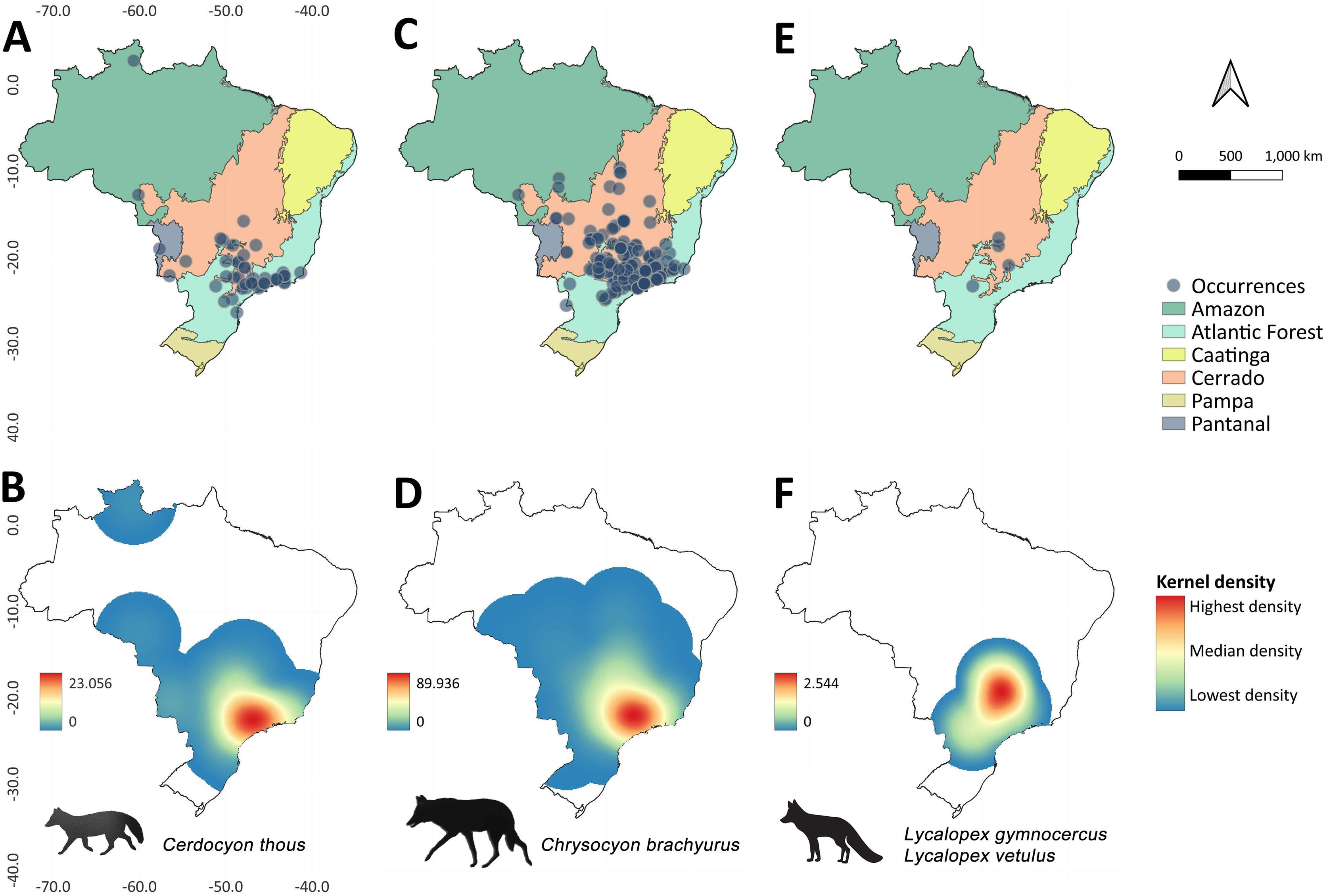
Occurrences of canids in urban areas throughout Brazil between 2010 and 2022, and the respective heatmaps based on Kernel approach showing spatial hotspots (highlighted in red) with high rates of occurrences. Canids species are: Cerdocyon thous (A-B), Chrysocyon brachyurus (C-D), Lycalopex gymnocercus and Lycalopex vetulus (E-F).
The GAM predicting occurrences of carnivorous mammals in urban areas of Brazil accounted for 65.8% of the total deviance (adjusted R2 = 0.658), and presented low overdispersion (dispersion parameter = 0.3287) suggesting that the model variance was well-adjusted to the observed data. The GAM model showed significant estimates mainly for HPD (edf = 2.79; χ2 = 19.74; p < 0.001, Table 1; nonlinear effect range approximately −0.10 to 0.50; Fig. 4). Additionally, other variables were statistically significant predictors of the variation in occurrences of carnivorans in Brazilian urban areas. For instance, the extension of roads (edf = 3.71; χ2 = 18.27; p < 0.001, Table 1; nonlinear effect range approximately −0.05 to 1.50; Fig. 4), HDI (edf = 2.64; χ2 = 14.77; p < 0.001, Table 1; nonlinear effect range approximately −0.05 to 1.10; Fig. 4), municipality size (edf = 0.9; χ2 = 8.62; P < 0.001, Table 1; linear effect range approximately −0.10 to 1.50; Fig. 4; Table 1), and elevation (edf = 0.78; χ2 = 3.32; p = 0.035, Table 1; linear effect range approximately −0.10 to 0.05; Fig. 4) were found to be important to predict these patterns. Additionally, the occurrences of carnivorans in Brazilian urban areas were marginally significant (p ≤ 0.10) explained by the poverty index (edf = 1.28; χ2 = 2.75; p = 0.09, Table 1; nonlinear effect range approximately 0.05 to 0.00; Fig. 4) and natural habitat (edf = 0.68; χ2 = 2.04; p < 0.07, Table 1).
Results of GAM in predicting 605 independent occurrences of mammalian carnivorans in urban areas throughout Brazil and, GAM models adjusted for each species with occurrence points >10. Significant (at p ≤ 0.05**) and marginally significant (at p ≤ 0.10*) variables are highlighted by asterisks and biome was used as a random effect. Acronyms were: HPD: population density; Roads: extension of roads; HDI: human development index; Size: municipality size; POV: poverty index; Lights: nocturnal lights hotspots; PA proximity: proximity of any protected area; Veg height: vegetation height; Biomass AG: biomass above ground; edf: effective degrees of freedom; and df: degrees of freedom.
| Response | Distrib. and link | Predictive variable | Edf | df | χ2 | p-value | R2 (adj.) | Deviance explained |
|---|---|---|---|---|---|---|---|---|
| Total number events | Poisson(log) | s(HPD)** | 2.79 | 9 | 19.74 | <0.001 | 0.658 | 63.8 |
| s(Roads)** | 3.71 | 9 | 18.27 | <0.001 | ||||
| s(HDI)** | 2.64 | 9 | 14.77 | <0.001 | ||||
| s(Size)** | 8.96 | 9 | 8.62 | <0.001 | ||||
| s(Elevation)** | 7.78 | 9 | 3.32 | 0.035 | ||||
| s(POV)* | 1.27 | 9 | 2.75 | 0.091 | ||||
| s(Natural_habitat)* | 6.80 | 9 | 2.04 | 0.072 | ||||
| s(Lights) | 8.58 | 9 | 0.00 | 0.715 | ||||
| s(PA proximity) | 2.67 | 9 | 0.00 | 0.530 | ||||
| s(Veg height) | 2.45 | 9 | 0.00 | 0.670 | ||||
| s(Biomass AG) | 1.74 | 9 | 0.00 | 0.839 | ||||
| s(Defaunation) | 3.86 | 9 | 0.63 | 0.191 | ||||
| Puma concolor events | ||||||||
| Poisson(log) | s(Size)** | 0.73 | 9 | 2.78 | 0.044 | 0.17 | 23.4 | |
| s(POV)* | 0.91 | 9 | 2.86 | 0.046 | ||||
| s(HPD)* | 1.12 | 9 | 2.89 | 0.053 | ||||
| Chrysocyon brachyurus events | ||||||||
| Poisson(log) | s(HPD)*** | 2.23 | 9 | 11.23 | <0.001 | 0.53 | 55.1 | |
| s(Size)* | 7.70 | 9 | 3.38 | 0.019 | ||||
| s(Roads)* | 2.08 | 9 | 5.57 | 0.029 | ||||
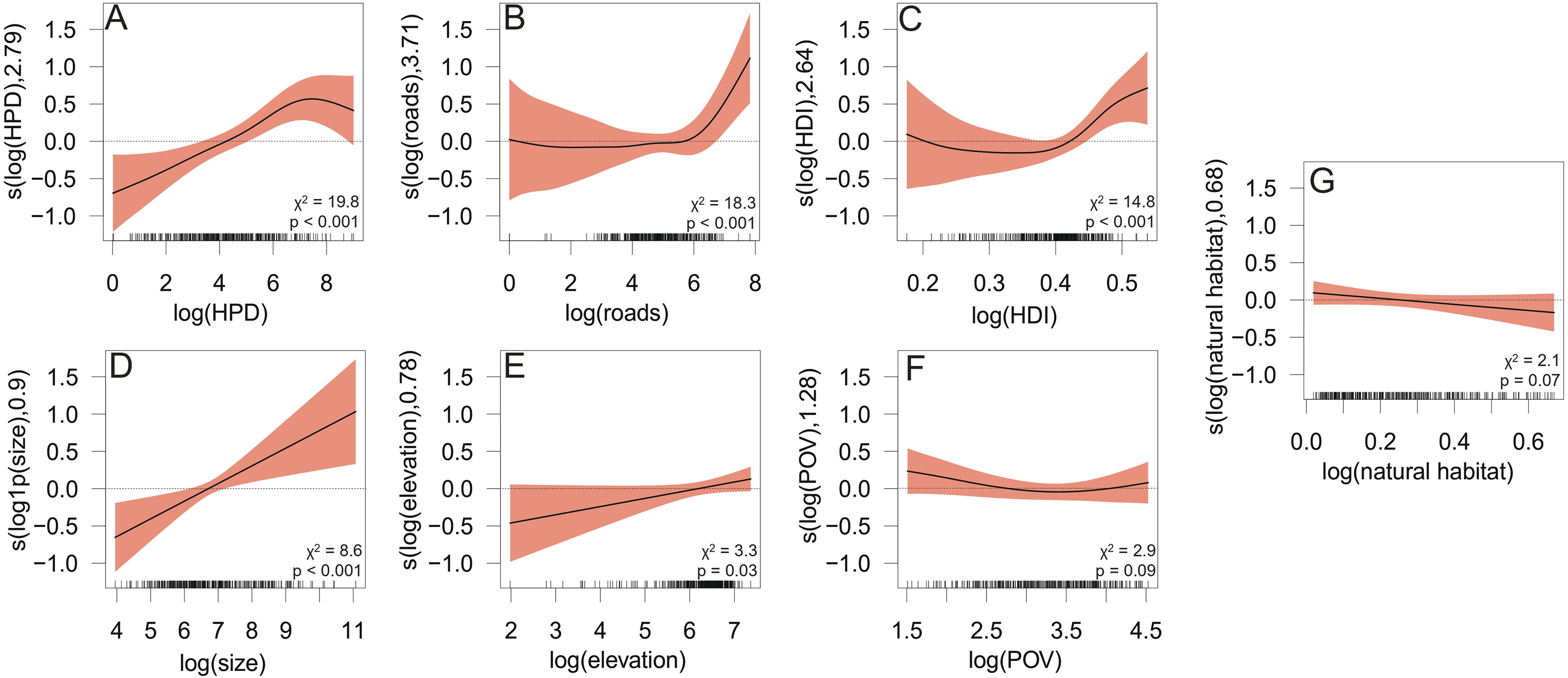
Significant (A to E; p ≤ 0.05) and marginally significant (F and G; p ≤ 0.10) effects (and its confidence intervals of 95%; shaded-coral) derived from a generalized additive model (GAM) to predict the occurrences of mammalian carnivorans in urban areas throughout Brazil. The x-axis scales are based on the log-transformed data, while the y-axis scales represent the smoothed (s) effect size of predictive variables.
The GAM model for P. concolor (puma) showed significant estimates, evolving mainly municipality size (edf = 0.73; χ² = 2.78; p = 0.04; Table 1; linear effect range approximately −0.5 to 0.8; Fig. S2; Table 1). Additionally, the poverty index (edf = 0.91; χ² = 2.86; p = 0.04; Table 1; nonlinear effect range approximately −0.05 to 0.05; Fig. S2; Table 1), and HPD (edf = 1.12; χ² = 2.89; p < 0.05; Table 1; nonlinear effect range approximately −0.4 to 0.5; Fig. S2) were statistically significant predictors of the variation in occurrences of puma in Brazilian urban areas. The GAM model for C. brachyurus also showed significant estimates, including HPD (edf = 2.23; χ² = 11.23; p < 0.001; Table 1; linear effect range approximately −1.4 to 0.8; Fig. S3; Table 1). Additionally, municipality size (edf = 7.70; χ² = 3.38; p = 0.01; Table 1; linear effect range approximately −1.3 to 1.4; Fig. S3; Table 1), and the extension of roads (edf = 2.08; χ² = 5.57; p = 0.03; Table 1; nonlinear effect range approximately −0.9 to 1.5; Fig. S4) were statistically significant predictors of the variation in occurrences of maned wolf in Brazilian urban areas. For L. pardalis, P. onca, and C. thous (i.e., with sufficient occurrence points > 10), the GAM models did not pinpoint any significance for the predictor variables in relation to occurrence in urban areas, neither social, geophysical, or environmental variables (p > 0.05).
DiscussionThe chronic and rapid decrease of wilderness globally will make the interaction between wildlife and humans unavoidable, potentially increasing human-wildlife encounters and conflicts in both urban and peri-urban areas (McKinney, 2002). Our results clearly showed that the vast majority of mammalian carnivorans occurrences in urban environments were located in densely settled areas throughout Southeast Brazil, overlapping the southern portion of the Atlantic Forest and Cerrado biomes and covering an area of approximately 586,265 km2. These results precisely reflect the pattern of historic urban expansion across Brazil, where natural habitats were constantly replaced by agricultural and urban landscapes, causing massive rates of habitat loss and fragmentation and decreasing dramatically the regional diversity. Southeast Brazil is home to approximately 84.8 million inhabitants, representing 41.8% of the total Brazilian population (IBGE – Instituto Brasileiro de Geografia e Estatística, 2021), whereas their cities continue to expand outward, altering natural habitats. Against both rapid and long-term habitat loss across tropical landscapes, urban areas and its resources will assume greater importance in wildlife conservation (Bateman and Fleming, 2012), given that many animals will increase their interface with anthropogenic landscapes across their home ranges (Gehrt et al., 2010). Thus, human-wildlife conflicts pose a significant conservation challenge for wildlife, particularly in anthropized landscapes such as urban environments and surrounding areas (McKinney, 2002).
We have identified 10 species of medium- to large-bodied mammalian carnivorans belonging to the Canidae and Felidae families, which exhibit high levels of occurrence in urbanized landscapes across Brazil. Notably, 70% of these species are either included in global threat categories as defined by the IUCN (2023) or are listed under specific threat categories according to the Brazilian Red List (MMA, 2022). The transformations occurring within the physical environment during the Anthropocene, driven by factors such as the expansion of road infrastructure, urbanization, and habitat loss, pose significant threats to biodiversity (Young et al., 2016). These threats are of major concern and have the potential to lead to the local extinction of small, isolated populations of numerous carnivorous mammal species (Bateman and Fleming, 2012; Young et al., 2016). The substantial proportion of species falling within threat categories underscores the urgency for targeted conservation endeavors, policy interventions, and comprehensive public awareness movements. These collective actions are vital to counteract the adverse impacts resulting from anthropogenic activities upon any wildlife species.
Studies support that medium-bodied carnivorans, such as foxes (Vulpes vulpes) and coyotes (Canis latrans), consume a wide variety of prey and other resources in urban environments provided by humans (Gehrt et al., 2010; Bateman and Fleming, 2012; Magle et al., 2021). For instance, in southern California coyotes exhibit variations in their diet in response to “urban” and “suburban” environments (Larson et al., 2020). Specifically, the consumption of anthropogenic items increased proportionally with the high-level of urbanization (Larson et al., 2020). In Zürich, Switzerland, more than half of red fox stomach contents contained anthropogenic-based foods (see, Bateman and Fleming, 2012). However, a wide range of these studies is concentrated in temperate regions of the northern hemisphere. Our results provide the first support for understanding how large Neotropical carnivorans are enterprising over a new environmental configuration of global landscapes, which are increasingly adjacent to their native ranges. For example, pumas exhibited higher occurrence densities in several Brazilian biomes. Studies suggest a greater plasticity of this species in exploring altered environments in search of resources such as shelter and food (Mazzolli, 2010). However, most of the studies have analyzed environments modified by agriculture and monocultures, with little attention given to urban environments (e.g., Mazzolli, 2010; Magioli et al., 2014). For the maned wolf — the second species most reported in urban environments — anthropogenic changes have resulted in range contraction in some areas and expansion in others, such as derived from Amazon savanisation (Coelho et al., 2018). Previous studies using niche models indicated that maned wolves tend to occupy locations farther from cities and roads (Coelho et al., 2018). However, in some situations, the species has been observed to occupy open habitats close to large urban centers (Coelho et al., 2008) and highly modified landscapes. Therefore, our findings can help to better understand the multiple factors influencing the persistence or extirpation of maned wolves in specific regions across Brazil. We highlight that there is still no theoretical support to classify any of this species recorded in this study as either urban adapters or urban avoiders (see McKinney, 2002). Further studies that consider the changes in natural history, ecological, and behavioral factors against modern challenges are of great importance.
Differently from our initial expectations, the environmental variables did not show any relationships with carnivorans occurrence across Brazilian urban areas. On the other hand, social and geophysical variables (i.e., HPD extension of roads, HDI, size, and elevation) were significantly associated with their occurrences in urbanized areas. We observed a consistent pattern when generating single models for the species documented in this study. However, it is noteworthy that only the models for pumas and maned wolves exhibited statistical significance for some of the variables, such as municipality size, roads and HPD (Table 1; Fig. S1 and S2). However, this pattern contradicts the expected biodiversity responses to anthropogenic threats, which typically reduce species diversity, particularly in the temperate realm (Sanderson et al., 2002; Bateman and Fleming, 2012; Bruskotter et al., 2017; Magle et al., 2021).
The high significance of HPD as an important predictor variable indicates that human density reflects regions where these carnivores had their habitat contracted and, therefore frequently appearing in urban areas. Higher human densities lead to higher levels of influence on nature. The consequences of interactions between human population density and the environment depend on the nature of the interaction and the particular species, ecosystems, or processes considered (Sanderson et al., 2002). The human population density, one of the components of the Human Footprint Index (HFI), provides a comprehensive view of human impacts on ecosystems and can be used to quantify habitat degradation (Sanderson et al., 2002). For example, mammalian movements in areas with a comparatively high HFI represented, on average, half to one-third of the extent of their movements in areas with a low human footprint, likely owing to changed behaviours and physical limitations (Tucker et al., 2018). Additionally, considering the HDI associated with the HPD could offer a more holistic understanding of how human activities affect carnivorans occurrences and habitat quality in urbanized regions. It is possible that the availability of food resources provided by human activities, along with access to urban areas, may be contributing to attracting these animals to human-altered environments. There are several cases of wildlife occupying urban environments worldwide, such as some medium-bodied omnivorans (especially raccoons (Procyon lotor) and opossums (Didelphis virginianus)) and carnivorans (foxes (V. vulpes) and coyotes (C. latrans)) that forage in garbage, vegetable gardens, and consume a wide variety of prey in urban environments (Gehrt et al., 2010; Bateman and Fleming, 2012; Larson et al., 2020).
Besides predictions derived from HPD and HDI, we identified municipal size and elevation as important factors in predicting carnivorans occurrences. These variables may be related to socio-economic aspects, such as available infrastructure and the level of urbanization, which can affect resource availability and the tolerance of mammals towards urban areas (Scott et al., 2014). Furthermore, we found a strong positive nonlinear effect range of the extent of roads in the occurrences of carnivorans in urban environments. Nonetheless, we suggest that within our model, the “extent of roads” variable is partially linked with both municipality size and HDI. This assertion stems from the observation that in more affluent economies, particularly in major urban hubs, infrastructure elements such as road networks exhibit significantly superior scale compared to the countryside. The construction of roads can alter habitats, either by providing access to new areas or creating physical barriers that limit the movement of some particular species (Pinto et al., 2022). The presence and impact of roads in urban environments significantly influence the distribution and survival of mammalian carnivorans, making it essential to mitigate the negative effects on wildlife in response to the expansion of urban landscapes over natural habitats (Pinto et al., 2022).
Although the relationship of poverty index and natural habitat with the occurrences of carnivorous mammals in urban areas were only marginally significant, their influences may still be relevant. It is widely accepted that biodiversity loss and poverty are linked problems and that conservation and poverty reduction should be tackled together (Adams et al., 2004). The marginal significance observed suggests the need for further investigation and a more nuanced exploration of potential connections. The intricate interplay between socio-economic factors, represented by the poverty index, and ecological variables, such as natural habitat availability, could contribute to a more comprehensive understanding of the factors shaping carnivorous mammal distribution in urban landscapes.
Our findings can contribute to understand the magnitude of mammalian carnivorans presence across urban environments in Brazil. Nevertheless, we acknowledge that our findings may only capture a fraction of occurrences in urban areas throughout the country. For example, conflicts between humans and wildlife in marginalized communities and impoverished areas may be significantly underreported, as a substantial portion of the country lacks local news outlets or newspapers, resulting in information being neglected. These results shed light on the potential increase in human-wildlife encounters and conflicts in urban environments (e.g., roadkills due to greater vehicle flow; Pinto et al., 2022). Further, our results pinpoint the higher odds of unexpected encounters with humans, concerns regarding the cross-transmission of zoonoses, and predation of domesticated animals (Bateman and Fleming, 2012). Human-wildlife conflicts represent an important socio-ecological dimension for both wildlife and humans, given that conflicts are real or perceived negative biological, economic, social, or political interactions (see Frank et al., 2019).
Finally, our results provide important insights into the intricate interplay of social and geophysical variables influencing the presence of carnivorans in urban environments in Brazil. We can recommend, for instance, that large cities with high population density need to invest in Wild Animal Triage Centers (CETAS), training professionals for wildlife management and that therefore, can effectively act as hubs for smaller neighbours cities. This issue is particularly crucial given that these urban areas have exhibited the highest occurrence rates in recent years. Furthermore, delving into studies focused on unravelling the adaptive mechanisms of these carnivorans to urban environments across Brazil, exploring how shifts in urban characteristics might impact their populations, would indeed contribute for a more comprehensive understanding of these crescent phenomena.
Declaration of interestsThe authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
FundingJAB was supported by Conselho Nacional de Pesquisa e Desenvolvimento Científico e Tecnológico (CNPq) postdoctoral fellowship (grant 150261/2023-3).
Author contributionsADP: conceptualization, data acquisition, data analysis and figures, writing & revising the original draft; NCI and KLK: conceptualization, data acquisition; JAB: conceptualization, data analysis, review and major editing.
The following is Supplementary data to this article:


