





Restoring tropical forests still relies on expensive tree planting. Timber production from native trees offers a promising opportunity to make restoration financially viable, but species growth data are lacking. We assessed the potential of tropical forest restoration plantations for producing native timber in the Atlantic Forest. For that we inventoried a chronosequence of unmanaged restoration plantation sites with ten commercial native tree species. Then we developed growth models and used the Growth-Oriented Logging (GOL) method to inform targeted management decisions, including an optimized timber-focused scenario, based on growth and bole quality assessment. Usually, growth-rate classes for saw wood production would be defined according to the time necessary for achieving 35 cm in DBH. Harvesting age was markedly reduced using GOL for species of fast-growth (from <50 to <25 years), intermediate-growth (from 50 to 70 to 25–50 years), slow-growth (from >70 to 50–75 years). Following GOL, basal area was 42 ± 30% larger at harvest (taking 35 ± 23% more time) or 60 ± 16.5% smaller (taking 66 ± 16% less time). The optimized scenario speeded tree-size achievement and anticipated harvesting in an average of 58 years. Species selection, individualization, and silvicultural management are critical for producing timber in forest restoration programs.
The global demand and enthusiasm for forest restoration has never been so high (Sacco et al., 2021). The world is witnessing a rally of forest restoration and tree planting pledges, with multi-million hectares and trillion-trees campaigns (Holl and Brancalion, 2020; Goymer, 2018). But implementing large-scale, long-lasting forest restoration requires more than aspirations; it must overcome critical and unresolved challenges that have constrained restoration programs worldwide (Fagan et al., 2020; Reid et al., 2018). One of the main challenges is its financial viability. Forest restoration relies, in most cases, on the reconversion of agropastoral land uses back into forests and requires large investments to kickstart forest succession in severely degraded sites through tree-planting approaches (Holl and Aide, 2011). Incentive mechanisms, such as payments for ecosystem services, may not be enough to overcome land opportunity, implementation, and maintenance costs. A promising opportunity to make active restoration financially viable is the production of native tree species timber. Still, the lack of reliable growth data for most species has been a central limitation for timber-oriented restoration projects.
Biodiverse plantations of native species have been primarily established with the main goal of ecologically restoring native forests, maximizing habitat for multiple species (Despot-Belmonte et al., 2017), soil protection (Deng et al., 2012; Krainovic et al., 2020), hydrological services (Ahmad et al., 2001), climate change mitigation (Bastin et al., 2019; Griscom et al., 2017), socioeconomic benefits (Erbaugh and Oldekop, 2018), and legal compliance (Rodrigues et al., 2009). Conversely, the illegal exploitation of forest remnants, along with commercial monocultures of exotic species, has been meeting a portion of the market demand for timber (Brancalion et al., 2018). In Brazil, highly diverse restoration plantations have been historically established on private lands, mainly as riparian buffers, to comply with the federal legislation, whereas commercial tree plantations (9.55 Mha) are dominated by eucalypt (7.47 Mha, 78%), and pine (1.70 Mha, 18%) stands. The few commercial native tree species cultivated largely for timber in their original domain are Araucaria angustifolia (Bert.) Küntze and Schizolobium parahyba var. amazonicum (Huber ex Ducke) Barneby, which occupy, together with other native species, only 3.4% of the tree plantation area (Brazilian tree industry, 2021).
Native and exotic timber production has occurred in contrasting conditions. Whereas native timber exploitation has been concentrated in large tracts of old-growth forests in the Amazon (Brancalion et al., 2018), based on the harvesting of old, huge trees, exotic timber production has predominated in short-rotation industrial plantations located in southern Brazil, in highly degraded and fragmented landscapes within Atlantic Forest and Cerrado areas (Gonçalves et al., 2013). The timber-derived products correspond to ∼90% of Brazil's total forest market value (Brazilian tree industry, 2021). The wood exploited from native forests yielded 518 million dollars in 2020 (IBGE, 2020), while those exploited from commercial plantations yielded 3.48 billion dollars. Improving native timber and non-timber production in restoration plantations can leverage large-scale restoration (Hua et al., 2022; Lamb et al., 2005).
The competition with illegal logging has always been a severe limitation for native timber production in commercial plantations. Still, the growing legal demands for restoring degraded lands in the country may change this scenario. Under the Native Vegetation Protection Law, farmers are required to maintain a minimum of 20%, 50%, or 80% of their landholding covered with native ecosystems, depending on the region. If they do not comply with an area of Legal Reserve (LR), they must compensate it off-farm or restore it locally. Harvesting timber through low-impact logging in the LR is allowed, although this is not possible in the Atlantic Forest domain (Brasil, 2006). As the deficit of LR encompasses several million hectares (Freitas et al., 2017; Metzger et al., 2019), many landowners are obliged to restore native forests, but ideally, they may count on native timber production to offset costs and make attractive profits.
Many emblematic species with high market value have been historically exploited in natural areas and now rely on plantations to be commercialized again, but the viability of timber production in restoration projects requires growing trees of market-valued timber in a reasonable time, with straight boles and a suitable diameter for a sawmill. However, little is known about the growth potential of native timber species, especially in biodiverse restoration plantations, which are not adequately managed for timber production (e.g., use of improved genetic materials, thinning and pruning, pest- and pathogen-control). Understanding the potential of restoration plantations to produce marketable native timber is a critical step for offsetting restoration costs and transforming it into a profitable activity. Given that restoration plantations are established with multiple native tree species and usually have an abundant regeneration of native colonizing species in the understory a few years after plantation, we expect that harvesting a share of the planted species could delay, but not hamper restoration trajectory over time. The level of impact will be affected by the harvested volume, harvesting method, and site features (e.g., slope, rainfall), so integrating timber production with forest restoration relies on careful planning and monitoring.
Growth predictions are essential prerequisites for guiding silvicultural management and harvesting decisions. Forest restoration growth models represent an important tool to assess critical aspects of timber production sustainability (Günter et al., 2011), revealing opportunities to guide management interventions and reduce the pressure of exploitation on native forests.
In this study, we aimed to understand the potential for native timber production in tropical forest restoration plantations. More specifically, we built growth models, defined harvest time, and designed an optimized scenario for timber production of native tree species introduced in biodiverse restoration plantations. We also evaluated the bole quality for saw wood production.
MethodsStudy sites and sampled speciesOur study was performed on 13 sites distributed across São Paulo State, southeastern Brazil (Figure and Table S1). Restoration sites were established by planting a diverse mixture of native tree species (30–100 species), aiming at ecological achievements. We selected areas with different ages (6–96 years old) to create a chronosequence representing the potential growth performance of the targeted timber species.
We selected 10 native timber species with different wood densities, historically overexploited for timber production. Most of these species are protected by law and no longer sold legally in the market, as they are endemic to the Atlantic Forest and Cerrado and have no viable commercial stocks available in natural areas. Yet, some of them (e.g., Hymenea courbaril L. and Handroanthus impetiginosus Mattos) are still exploited in the Amazon. For this study, we chose 10 species: Balfourodendron riedelianum (Engl.) Engl., Cariniana legalis (Mart.) Kuntze, Cedrela fissilis Vell., Centrolobium tomentosum Guillem. ex Benth, Esenbeckia leiocarpa Engl., Hymenea courbaril L., and Peltophorum dubium (Spreng.) Taub. which are characteristic from the mid-successional stage (CONAMA, 1994), and, Handroanthus impetiginosus Mattos, Astronium graveolens Jacq. and Myroxylon peruiferum L.f. which are typical from late-successional stages (Balestrin et al., 2019; Ribeiro et al., 2005).
We measured all individuals from these species at the study sites (Diameter at Breast Height - DBH – 1.30 m above the ground) and classified their bole quality into (1) straight stem (best quality); (2) straight stem with a small number of branched or crooked and unbranched stem (high quality); (3) crooked and unbranched stem (low quality); (4) crooked and branched stem (worst quality - non-commercial). We sampled 172.5 (average; SD = 69; min-max = 77–314) individuals of each species (Table S2) and a total of 1725 trees. Due to variations of species abundance in our sites, it was not possible to obtain a balanced sample, with the same number of individuals per species per site. We did our best to select sites with similar biophysical conditions (soil type – Fig. S1, climate, and vegetation type) and restoration management (diverse plantations, 3 × 2 m spacing and, no silvicultural management). However, we recognize that uncontrolled local variations in site conditions, and management and tree density co-vary with age and this could not be controlled/identified in our analysis, as we have only one sample per age. Consequently, it is important to recognize this important limitation of our study.
Data analysisTo determine the best-fit growth model, we tested the Asymptotic, Four-Parameter Logistic model, Michaelis-Menten, and variations of Logistic models for each species, totalizing eight different models. As tree growth is a nonlinear process, the non-linear models are expected to outperform well (Chen et al., 2022). The fit statistics used to judge the performance of the model were the proportion of variance explained (R2), relative standard error (RSE%), the mean absolute percent error (|E|%), and also the evaluation of the residuals distribution, degree of adjustment between observed and estimated values through the Akaike Information Criterion values (AIC index) and the significance of the coefficients (Vaughan et al., 2021). The RSE% and the determination of the nonlinear (weighted) least-squares estimates of the parameters of the nonlinear models and the significance of the coefficients were calculated following the Gauss-Newton algorithm using a nonlinear function (nls) from the R software in version 2022.12.0 of the RStudio (R Core Team, 2021). After the initial tests, we modeled DBH growth and basal area of selected species across the chronosequence using the calculated equivalent diameter (diameter of the commercial stem for bifurcated trees) on the Michaelis-Menten model (Coates and Burton, 1999; Evans et al., 2015; Vaughan et al., 2021) which was the model that yielded the best outcomes for all species. We used descriptive statistics to quantify bole quality for each species.
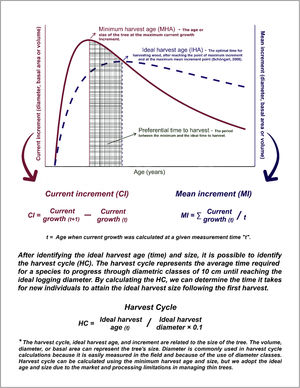
Analysis of tree growthUsing the DBH estimated for each age to provide a time series analysis on tree age and diametric and basal area increments, species were classified in fast-, intermediate-, slow-, and super slow-growth, based on the time needed for reaching a 35 cm DBH – a DBH size traditionally employed to decide on harvesting for timber in sawmills (standard approach) and a second classification was based on the time recommended by the Growth-Oriented-Logging concept (GOL approach). The diameter of different tree species, from the youngest to the oldest restoration site, was used to calculate the basal area increments that were accumulated to build individual growth curves (Brienen and Zuidema, 2007). The mean cumulative growth curve describes the relationship between tree age and the size of a given species (Resende et al., 2020; Schöngart, 2008; Stahle et al., 1999; Worbes and Schöngart, 2019), which, together with the cumulative increment curve allows the definition of the best productivity versus time. The GOL approach was developed for volume growth, but given the lack of volumetric equations for the studied tree species, we used basal area growth as an alternative. The conceptual GOL approach and the rationale for calculating current increment (CI), mean increment (MI), minimum harvest diameter (MHD), ideal harvest age (IHA), and wood harvest cycle (HC) is described in Box 1. Then, we employed the GOL concept to perform a growth analysis to compare the ideal harvest time and tree size with the 35 cm DBH threshold and understand if it is a good reference metric for deciding on harvesting and evaluating if it reflects or not a good relationship between time and productivity.
, the Mean Increment (MI), and the calculation and meaning of the Harvest Cycle (HC).")
Finally, we built productivity scenarios using the 30% highest DBH values found for each species per site/age. The adoption of such growth for modeling the stands resulted in a reduction of 53% (±14) in the time needed for reaching a 35 cm DBH, representing a 2-fold gain within the same period. Such reduction is realistic given that several studies showed an improvement of up to 7-fold in diameter, volume, or biomass ("size of the tree") growth after the adoption of silvicultural treatments or minimum improvement programs at the species-level. We used this potential growth to calculate the GOL, considerate possible if plantations had ideal silvicultural treatments such as improved soil preparation, knowledge of plant nutritional requirements, fertilization and weed control (Barreiros et al., 2007; Brancalion et al., 2019; Campoe et al., 2014; Ferez et al., 2015; Stape et al., 2010), selection of seed trees and appropriate seed collection methods, improved vegetative propagation methods, seedling improvement (Harwood et al., 2015; Oliveira et al., 2019; Rolim et al., 2019), pruning, aboveground biomass management, selective thinning, fire control and other (Amaral et al., 2019; De Graaf et al., 1999; Eufrade-Junior et al., 2021; Gonçalves et al., 2004; Zhang et al., 2012). The difference between the standard growth model (all data) and the model built for ideal conditions (30% largest trees) was confronted with the estimation of potential of wood production.
ResultsHarvesting time and diameterBased on the time required for achieving a DBH of 35 cm for harvesting, the species were classified as having fast (<50 years), intermediate (50–70 years), and slow growth (>70 years; Fig. 1). When the GOL approach was used, species were grouped into four growth-rate classes (fast: <25 years; intermediate: 25–50 years; slow: 50–75 years; and super slow: 75–100 years; Fig. 2). The GOL approach recommended shorter harvest cycles of thinner boles for increasing timber productivity/time relation for most species. For recommendations of a faster first harvest, the approach resulted in an average harvest time 66 ± 16% shorter and a basal area 60 ± 16.5% smaller, whereas the recommendation of delayed harvest yielded a 35 ± 23.4% longer time and a basal area 42 ± 30% greater (Table 1). The GOL curve for each species is shown in Figure S3.
. The vertical dashed lines indicate the age class in which a DBH of 35 cm is reached.")
.")
Recommended age and DBH for logging resulted from using the growth-oriented-logging approach and the fixed DBH of 35 cm (standard approach). IBA: Ideal Basal Area predicted by the GOL approach; BA is the Basal Area for each approach. BA35 is the basal area when DHB is 35 cm (962.1 cm2).
| Species | GOL approach | Standard approach | Variation GOL/Standard (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Age (yrs.) | BA (cm2) | DBH GOL (cm) | Age (yrs.) | BA (cm2) | DBH (cm) | Age (yrs.) | BA (cm2) | DBH (cm) | |
| A. graveolens | 57 | 622.9 | 28.2 | 94 | 962.1 | 35 | 60.6 | 64.7 | 80.5 |
| B. riedelianum | 17 | 266.3 | 18.4 | <100 | 962.1 | 35 | <17 | 27.7 | 52.6 |
| C. legalis | 45 | 1208. 9 | 39.2 | 37 | 962.1 | 35 | 121.6 | 125.6 | 112.1 |
| C. fissilis | 92 | 1765.4 | 47.4 | 54 | 962.1 | 35 | 170.4 | 183.5 | 135.5 |
| C. tomentosum | 68 | 1004.0 | 35.7 | 67 | 962.1 | 35 | 101.5 | 104.3 | 102.1 |
| E. leiocarpa | 15 | 164.4 | 14.5 | <100 | 962.1 | 35 | <15 | 17.1 | 41.3 |
| H. impetiginosus | 86 | 1494.7 | 43.6 | 58 | 962.1 | 35 | 148.3 | 155.4 | 124.6 |
| H. courbaril | 25 | 271.9 | 18.6 | <100 | 962.1 | 35 | <25 | 28.3 | 53.2 |
| M. peruiferum | 41 | 486.5 | 24.9 | <100 | 962.1 | 35 | <41 | 50.6 | 71.1 |
| P. dubium | 19 | 486.1 | 24.9 | 45 | 962.1 | 35 | 42.2 | 50.5 | 71.1 |
The optimized scenario (using the 30% largest trees of each site) had the meantime for harvesting reduced by 25%, and the mean basal area increased by 38%. It represented an average anticipation of ∼13 ± 13 years in the ideal harvest age and a 48% increase in basal area (295 cm2/tree) (Fig. 3). As exceptions to this trend, C. legalis. and H. courbaril had their ideal time for harvesting prolonged, but the harvesting basal area increased by more than 50%, whereas C. fissilis had a 36.6% reduction in harvesting basal area (646.6 cm2/tree) but a 47 years anticipation in harvest time (51% faster than the GOL). Half of the species achieved a 35 cm DBH at the ideal harvesting age recommended by GOL in the optimized scenario, while 90% reached a 35 cm DBH before 60 years, except for one species of high wood density (E. leiocarpa Engl.) (Figure S3, Table S4, and S5).
Stem quality and harvest basal area (B) between the growth-oriented-logging models created using all sampled individuals and only the 30% largest individuals from each site.")
Most species had nearly half of their individuals with one stem and a quarter of them with two stems. The mean stem quality index was 1.82 ± 0.34. To all classes of stem number (1, 2, and 3), except for the multiple stems class (3+), 95% of the stems were classified as commercially viable (best, high, or low bole quality). The number of stems per tree remarkably increased in fast-growing species, once more than 75% of the trees in the other three categories had 1 or 2 stems. The occurrence of best and high timber quality was higher in trees with 1, 2, or 3 stems in all growth-rate classes. In all classes, 49.5% of the timber stock had only one stem, followed by two (24.2%), three (12.3%), and more than three (1.7%) stems (Figure S2).
DiscussionCritical questions related to the production of native timber in restoration plantations, like "Which species to plant?", "When can I harvest some trees?", "When does the payback begin? "How much should be invested?" and "Which yield to expect?" are still unsolved, undermining investments in this activity. We explored some of these questions using a chronosequence of commercially valuable native trees from restoration plantations. Most tree species analyzed have reasonable stem quality but slow growth, at least when compared to exotic species commonly used in the tropics, such as Australian Eucalyptus (Stape et al., 2010). We discuss alternatives to minimize some of these limitations and increase the viability of timber production in restoration plantations.
Most tropical trees valued for their timber are late-successional hardwood species, which grow slowly and only reach large sizes after decades. Their initial development in the forest understory, where light availability is lower and competition is high, contributes to concentrating biomass allocation to a single stem, thus resulting in boles of great quality for saw wood production. Such characteristics have favored the exploitation of old trees in forest remnants, allowing successive harvesting cycles (Schulze et al., 2008). Restoration plantations offer contrasting conditions for timber production. Tree seedlings are planted in open areas and have high light availability during their initial development, which favors the production of multiple stems. Such conditions could also favor tree growth, but late-successional timber species compete with pioneer trees in the early stages of restoration development, severely limiting their growth (Brancalion et al., 2020; Holl and Brancalion, 2020).
Thinning and pruning would help resolve these limitations, but these techniques are not commonly used in biodiverse restoration plantations. Biodiverse stands require ecological and silvicultural knowledge about the species being cultivated for prescribing thinning and ensuring that trees receive adequate light for their full growth (Rolim et al., 2019), while experiments with well-known species have shown that the stand structure (e.g., spacing between trees), has a significant impact on wood growth (Stape et al., 2010). Tree growth limitations have been observed in reduced-impact logging operations in the Brazilian Amazon, where the slow growth of late-successional species planted using the enrichment technique has frustrated time expectations of sustainable timber management (Pinto et al., 2021). Thus, just planting commercially viable native trees in restoration areas and waiting does not seem a viable option for efficient timber production. Timber production in restoration plantations is a novelty that requires further studies and advances to become a competitive land use.
Unlocking the high-quality timber potential of native tree species may require a robust research and development program to resolve critical scientific and technological barriers. Using the GOL approach as an alternative to adopting a pre-defined DBH for deciding when to harvest trees in a species-specific manner can greatly improve the silvicultural performance of restoration plantations for timber production. Managing trees based solely on their DBH is ecologically arbitrary since it ignores the need of each species to achieve their maximum productivity for the first harvest and to advance size classes during harvest cycles. Furthermore, the decision to harvest trees based on a fixed DBH could make forest restoration less attractive financially once trees must be maintained despite not growing well. Our results emphasize the need for species-specific growth rate data to optimize the management of restored forests. This ensures a balanced approach between time for harvesting and productivity, leading to reduced harvesting time for similar-sized plants during the first harvest and subsequent management periods. Furthermore, it results in trees of larger diameter within an equivalent growth period.
It is important to recognize, however, that using the GOL approach in restoration, in general, may favor harvesting trees of smaller DBH, which has important consequences for wood processing and quality. Thinner logs has reduced sawing yield and may be composed of a lower proportion of heartwood, which have higher market value due to the greater mechanical stability and resistance to predators and natural hazards (Chave et al., 2009). For example, the teak obtained from plantations in central Brazil has a high proportion of whitish sapwood, as opposed to the dark brown heartwood of larger native teaks exploited from forest remnants in Asia (Carmo et al., 2020; Pereira et al., 2021). Companies have sold whitish teak timber for a lower price but are finding alternative ways to use the timber commercially, considering its particularities (Lima et al., 2021). The regular wood mills that operate using timber exploited from native remnants in the Amazon, as well as those using large logs of eucalypt and pine produced in southern Brazil, may not be adequate to process small native trees coming from restoration plantations. Modern, more sophisticated equipment is required, and novel wood products and uses must be developed to better incorporate the native timber from restoration sites in the market. Recent advances in wood engineering, particularly in processing technology and log utilization rates, hold the potential to significantly broaden the possibilities of using this type of wood for diverse applications. The timber produced in restoration plantations may represent a potential shift from buying timber from unreliable sources, often illegal logging, to a controlled source based on tropical forest restoration.
The superior silvicultural performance of exotic species is a direct consequence of decades of research, development, and innovation performed by public and private agents (Gonçalves et al., 2013). The native tree species investigated here were not submitted to any level of silvicultural improvement and, as expected, still offer a large room for improvement opportunities in forestry sciences. Many valuable timber species historically exploited may not be good candidates for timber production as they grow too slowly. Restoration projects should consider intercropping slow-growing high-quality timber species with species of fast to intermediate growth to offset costs and anticipate revenues (Brancalion et al., 2020; Holl and Aide, 2011). Besides, the production of native timber in restoration plantations will be riskier than using exotic species from industrial monocultures due to the lack of clarity in the business plan (growth rate, payback, future price, and others) and market trend information. For this reason, business-as-usual decisions may not enable the implementation of this system, and more ecological studies about interactions between slow- and fast-growing species from different successional groups are needed to understand species performances and reduce risks. It is critical to consider that restoration plantations will deliver several other benefits, especially those related to biodiversity, soil, and hydrological aspects (Hua et al., 2022), non-timber forest products and other ecosystem services which could also be monetized and "bundled" with timber to offer attractive cash-flow to farmers.
ConclusionsA good perspective exists to achieve sustainable native timber management in forest restoration programs while providing ecosystem services. To achieve high productivity, species-specific criteria related to tree growth, the performance of combined tree species, management, and harvesting plans, as well as silvicultural treatments and research development and innovation must all be incorporated into the forest restoration value chain.
Conflicts of interestPHSB and RRR are partners of the Re.green company.
Conflict of interestsThe authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper. ☒ The authors declare the following financial interests/personal relationships which may be considered as potential competing interests:
Pedro Medrado Krainovic reports State of Sao Paulo Research Foundation provided financial support. Pedro Medrado Krainovic reports The Nature Conservancy provided financial support. Angélica Faria de Resende reports the State of Sao Paulo Research Foundation provided financial support. Catherine Torres de Almeida reports that the State of Sao Paulo Research Foundation provided financial support. Danilo Roberti Alves de Almeida reports the State of Sao Paulo Research Foundation provided financial support. Pedro Medrado Krainovic reports World Resources Institute provided financial support. Pedro H. S. Brancalion reports The Nature Conservancy provided financial support.
P.M.K. thanks to the financial support from Luiz de Queiroz School of Agrarian Studies Foundation (FEALQ - #104004), The Nature Conservancy (TNC), the World Resources Institute (WRI) and the São Paulo Research Foundation (FAPESP#2022/07712-5). In addition, the São Paulo Research Foundation (FAPESP) is acknowledged for financial support (grants #2019/24049-5, #2020/06734-0, #2018/21338-3, and projects #2018/18416-2, and #2020/06694-8).
The following is Supplementary data to this article:

