



The loss or reduction of animal populations and consequent extinction of ecological interactions in Neotropical forests demand urgent conservation initiatives to reverse these trends. One of the rainforests with the highest levels of mammal defaunation is the Brazilian Atlantic Forest. Local mammalian extinctions in the biome were evaluated to set out priorities. Researchers, reserve managers and ex situ animal keepers throughout the Atlantic Forest were connected through a reintroduction network. From 2010 to 2017, we reintroduced two important seed dispersers, the red-humped agouti and the brown howler monkey, in Tijuca National Park, Rio de Janeiro, with other species on their way. We monitored the reintroduced populations regarding demography, spatial patterns, diet and their effect on ecological interactions. They interacted with several plant species, including large-seeded ones. We found 25 dung beetles’ species interacting with howlers’ feces. As TNP lacked medium and large sized frugivores, the increased dispersal can have a disproportional effect on forest regeneration. Among the main constraints for refaunation programs we pointed out delays to obtain environmental licenses, scarcity of source populations and difficulties regarding quarantine, release and monitoring of the animals. Refaunation has shown promise as a low-cost, effective way to restore ecological processes in defaunated Neotropical forests.
In the last years, there has been an increased awareness that we may be going through Earth's sixth extinction wave, comparable to the great mass extinctions of the geological past (Barnosky et al., 2011). This process started approximately 50,000 years ago, with the extinction of most of the world's megafauna following human dispersal around the globe (Araujo et al., 2017), and has become increasingly severe in recent times, including many local extinctions and drastic population declines, especially in the biodiversity-rich tropics (Johnson et al., 2017).
The loss or reduction of animal populations can lead to the loss of ecological interactions in ecosystems (Valiente-Banuet et al., 2015). Janzen (1974) was the first to point out that the extinction of ecological interactions can impair ecosystem functioning and services (Balvanera et al., 2006). In forests where seed dispersal has been disrupted, for example, piles of fruits and seeds rot on the ground, while the vegetation appears to be intact – the so-called “empty forest” syndrome (Redford, 1992). In an empty forest, the chances of a large seed to become a tree are slim (Kurten, 2013).
In the last decades, there has been an increased interest in rewilding, aiming to restore ecosystems by translocating ecologically or locally extinct species, or by replacing them. The use of proxies to replace species extinct for decades, centuries or even millennia has been increasingly debated (pleistocene rewilding, Donlan et al., 2005), but such management practices are risky and have been heavily criticized (Caro, 2007; Oliveira-Santos and Fernandez, 2010). A simpler approach that has been widely used is the reintroduction of a single species’ population in an area from where it was extirpated (IUCN/SSC, 2013). The best example in Brazil is the successful reintroduction of the golden lion tamarin (Leontopithecus rosalia) in part of its original range in northern Rio de Janeiro state (Kierulff et al., 2012). However, its effects on the restoration of ecological processes have seldom been evaluated (Seddon et al., 2014). Besides, isolated reintroduction initiatives may not be enough to restore ecosystem functioning shattered by widespread defaunation, as any species’ biology imposes limitations on the interactions it can rewire.
To address this problem, Oliveira-Santos and Fernandez (2010) proposed the idea of refaunation – the restoration of native faunas. Implicit in their proposal was the so-called “Columbian baseline”, the use of species present by the time of European colonization. This approach would be based in the sequential reintroduction of a set of recently extirpated animal species, allowing the restoration of ecological interactions with the extant fauna and flora. This is similar to “trophic rewilding” (Svenning et al., 2016), with the difference that the latter prioritizes the restoration of trophic interactions more than the fauna itself; we use the two terms as synonyms hereafter. To optimize the recovery of interactions and to allow subsequent recolonizations by other species, Galetti et al. (2017a) suggested a logical sequence of reintroductions that should be followed for the trophic rewilding of Neotropical forests.
The Atlantic Forest of Brazil, one of the World's richest biodiversity hotspots (Myers et al., 2000) but widely defaunated (Galetti et al., 2017b), is an obvious priority for refaunation. The ambitious idea of refaunating Atlantic Forest sites in Rio de Janeiro led to our first reintroduction in 2010, and to the creation of the REFAUNA Project in 2012. Our first goal was to map and evaluate local extinctions (extirpations) of mammals in the Atlantic Forest, and identify where refaunation initiatives would be most needed. We compared original and present distributions of all medium and large-sized mammals that occurred in the Atlantic Forest when the Portuguese colonization started in 1500, and estimated which proportion of its original distribution each species had lost. We then created a virtual reintroduction network, the REFAUNA Network (http://refauna.wixsite.com/site), to connect researchers, reserve managers and ex situ animal keepers throughout the Atlantic Forest biome. Besides, with the REFAUNA Project, we also aimed to carry out the reintroductions themselves, to restore ecological interactions. Our first refaunation target was Tijuca National Park (TNP), a widely defaunated Atlantic Forest reserve within Rio de Janeiro city. TNP suffered with deforestation due to agriculture, especially coffee and sugar cane plantations from the XVII to the XVIII century, together with hunting pressure. After a great effort of reforestation in XIX century (Dean, 1995), forest cover was restored, but the fauna remained impoverished. As Tijuca Forest is surrounded by an urbanized matrix, most species cannot naturally reoccupy the area. The history and isolation of TNP make it a suitable natural laboratory for rewilding. There we have reintroduced two species of mammals so far (the red-humped agouti Dasyprocta leporina and the howler monkey Alouatta guariba), with other reintroductions planned for the near future. Herein we present the procedures adopted and the success these reintroductions had in restoring ecological interactions. We conclude discussing the challenges we faced in our efforts in order to provide insights for future reintroduction and refaunation initiatives in Neotropical forests.
Atlantic Forest mammal lossesOnly 11.7–16% of the Atlantic Forest original cover still remains (Ribeiro et al., 2009). Increasing population pressure along history led to intense forest fragmentation, which, together with poaching and diseases, have caused many local extinctions of medium and large bodied vertebrates (Canale et al., 2012; Galetti et al., 2017a), resulting in the loss of an average of 88% (±SE 9.6) of their original (1500 A.D.) distribution. Most Atlantic Forest remnants have lost most of their medium and large mammals.
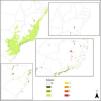
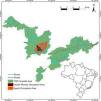
Bringing together the efforts of the actors involved: the REFAUNA NetworkOne of the main caveats of any reintroduction is to find a source population. We believe captive animals are usually the best choice because using wild-caught animals brings risks to wild populations, as mammals often occur in low densities. Through the REFAUNA Network we listed 474 ex situ animal keepers among zoos (120) and other breeding facilities (354). There was a lack of information regarding many animal sources, however, and only 71 of the listed keepers informed their stock sizes to the Brazilian environmental institute (IBAMA). Besides animal sources, we also listed 2910 researchers and other conservation agents, and 318 protected areas suitable for reintroductions or refaunation (Fig. 1). Only 14 protected areas had all the seven species analyzed on REFAUNA Network (see Table 1; A. guariba and Bradypus torquatus were analyzed only for Rio de Janeiro state) and did not need reintroduction programs (Fig. 1).
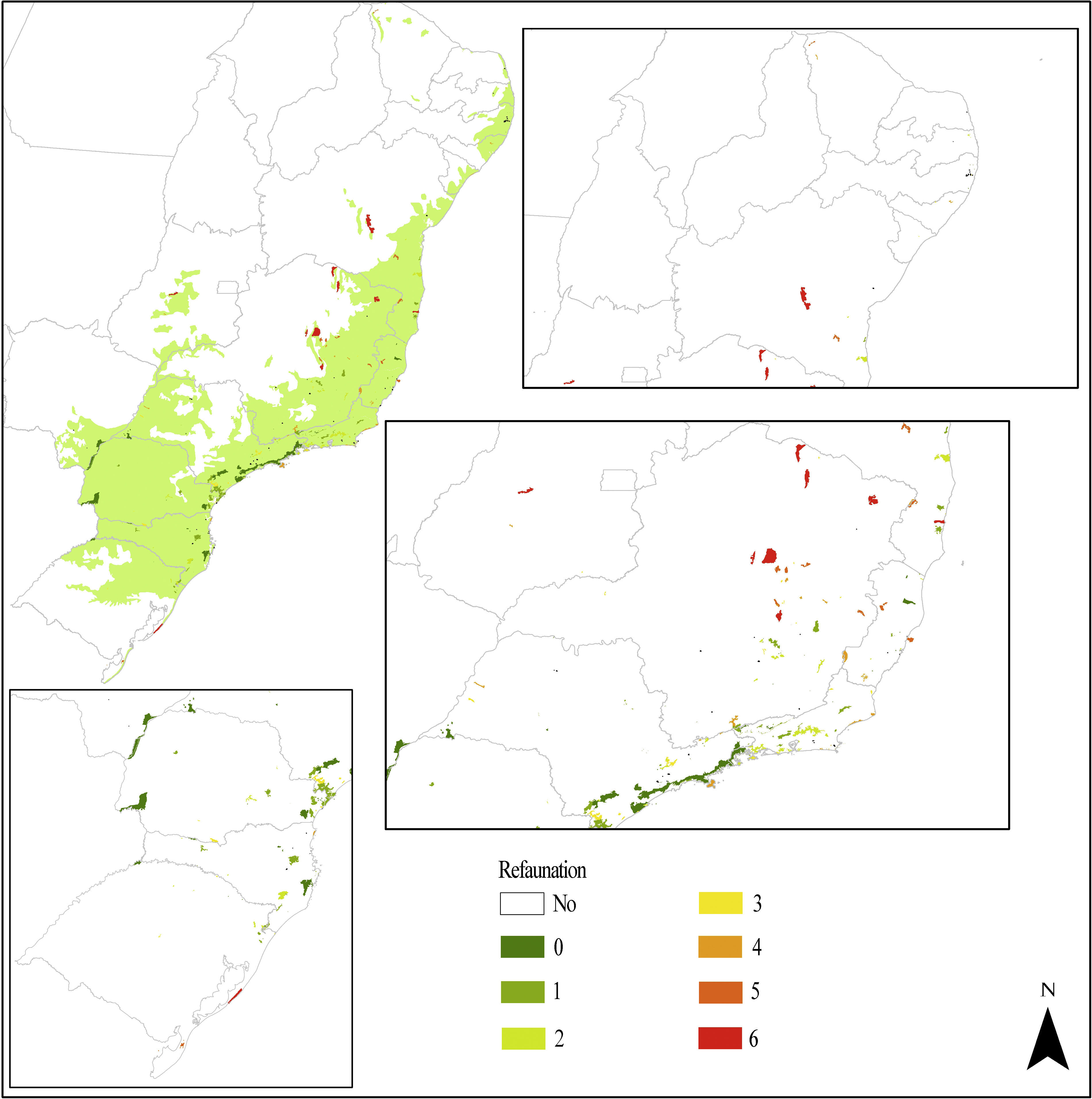
The 318 Atlantic Forest protected areas analyzed on REFAUNA Network. The color of protected area refers to the number of reintroduction programs needed for this unit. Species analyzed were: Agouti paca, Dasyprocta leporina, Tapirus terrestris, Pecari tajacu, Tayassu pecari, Leopardus pardalis and Panthera onca.
Mammalian candidates for refaunation in Rio de Janeiro state: reintroduction sequence based on Galetti et al. (2017), number of protected areas (PA) within each species’ geographical distribution in Rio de Janeiro state and number of those where the species actually occur. The last two columns show the number of ex situ animal keepers where each species is available (in Brazil as a whole), and total captive stocks for each species.
| Species | Reintroduction sequence | Number of suitable PA | Number of current PA where species occur | Number of ex situ animal keepers | Total animal captivity stock |
|---|---|---|---|---|---|
| Agouti paca | 1 | 24 | 18 | 30 | 724 |
| Alouatta guariba | 1 | 24 | 15 | 23 | 146 |
| Bradypus torquatus | 1 | 24 | 4 | 6 | NA |
| Dasyprocta leporina | 1 | 24 | 17 | 9 | NA |
| Tapirus terrestris | 1 | 6 | 0 | 44 | 146 |
| Pecari tajacu | 2 | 24 | 12 | 34 | 425 |
| Tayassu pecari | 2 | 31 | 4 | 35 | 1946 |
| Leopardus pardalis | 3 | 6 | 12 | 49 | 79 |
| Panthera onca | 4 | 0 | 0 | 35 | 92 |
An urgent target for the REFAUNA Network is Rio de Janeiro state, whose 26 fully protected areas (Biological Reserves and Parks) have lost most of their medium and large sized mammals (Table 1; Rocha et al., 2004). There are no resident populations of tapirs or jaguars in the whole state (Medici et al., 2012; Zeller, 2007). Although there is no single reserve large enough to maintain a viable jaguar population, the so-called Central Fluminense Mosaic covers nearly three hundred thousand hectares (Costa et al., 2010), and if its 29 protected areas are properly managed, it represents the only opportunity for reconstructing “complete” faunas, including apex predators.
Tijuca National Park as a laboratory for refaunationTijuca National Park (TNP) is a suitable laboratory for refaunation experiments. The area is a 3953ha forest fully embedded in a metropolitan matrix – which means that fauna released there can be easily controlled and could not disperse to other forest fragments. A previous attempt for restoring TNP's fauna was carried out from the late sixties by Coimbra-Filho et al. (1973). Among other animals, individuals of six mammalian species – some of them still present in the park – were released, including 32 agoutis (D. leporina). The releases were widely separated in space and time, the released individuals were not monitored, and the population of agoutis disappeared soon afterwards. Our first steps have been the reintroductions of the red-humped agouti (D. leporina) and the brown howler monkey (A. guariba). Agoutis and howlers were presumably extinct due to habitat loss (TNP was deforested for sugar cane and coffee plantations and reforested in the 19th century) and over-exploitation. Although domestic and feral dogs can prey upon native fauna and extirpate populations (Lessa et al., 2016), we do not believe that this was the main cause of agouti extinction in TNP. The habitat has been restored, and there are no signs of intensive poaching in TNP today. Therefore, we believe the causes for their original extinction are now controlled.
Agoutis should be part of the early stages of trophic rewilding because they occupy a low position in the trophic chain and because of their pivotal role in restoring ecological interactions (Galetti et al., 2017b). The species disperses large seeds through long distances (>100m; Jansen et al., 2012), and carry them toward locations with lower conspecific tree densities (Hirsch et al., 2013). These characteristics make seed dispersal by agoutis highly effective, enhancing seed survival.
Howler monkeys had been locally extinct for over a century. There were no large primates left at TNP and howlers can have an important role in restoring ecological processes (seed dispersal; nutrient cycling). They are folivore-frugivores with a close correlation with dung beetle (Scarabaeidae: Scarabaeinae) abundances (Culot et al., 2017), and the use of howlers’ dung by coprophagous beetles can enhance quality of micro-site seed deposition and help nutrient cycling and soil fertilization processes (Nichols et al., 2008). Hence, their role in restoring ecological services provided by extirpated species on defaunated areas, such as TNP, makes them suitable for reintroduction in early stages of refaunation (Galetti et al., 2017b).
First steps toward refaunation: results of reintroductionsThe source for agouti reintroduction was a semi-captive population at Campo de Santana, an urban park with a very high agouti density (about 50individuals/ha). The population was founded by an uncertain number of captive animals of the old Rio de Janeiro city Zoo in the early 1900s, whose original population was native to Rio de Janeiro state. Before translocation to TNP, agoutis went through a quarantine period to assure their health. Most individuals were released using a soft-release protocol, with two weeks of adaptation in an enclosure within TNP with food supplementation. Thirty-one agoutis were released from 2010 to 2014 (17 females and 14 males) and monitored by radio-tracking. Animals had high survival rates, quickly became independent from food supplementation and reproduced in the wild, configuring a short-term reintroduction success (Cid et al., 2014).
Population monitoring through mark-resighting yielded estimates of around 30 wild-born individuals in March 2015, with agouti numbers reaching as much as 41 individuals the year before. As overall population growth was positive, with all individuals being wild-born, the population was clearly able to grow without the aid of further releases. Thus, the reintroduced population has entered a growth stage, with a well-established distribution in the release area, and can be considered successful, at least in the medium-term (Fig. 2; Kenup et al., 2017).
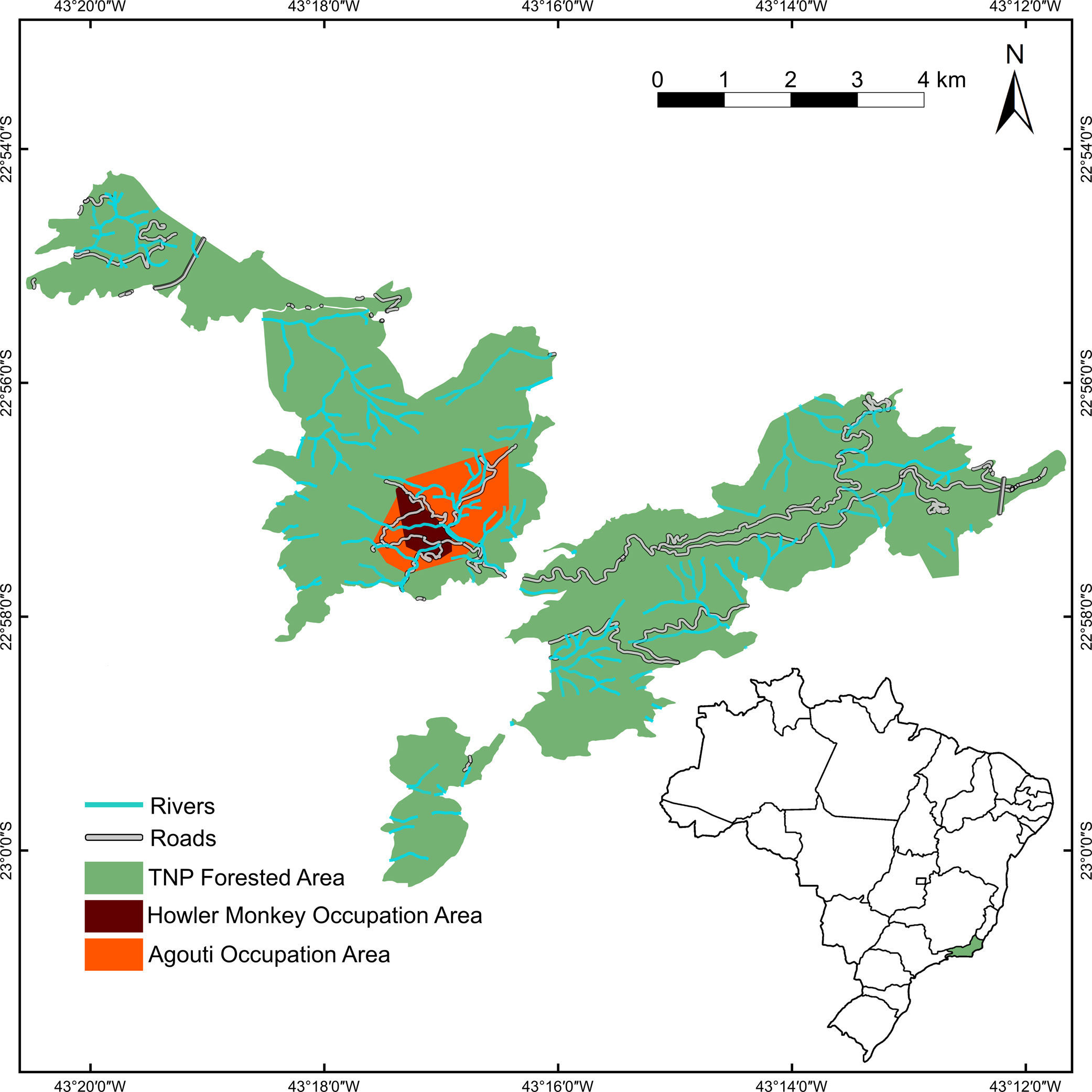
Location of the study area in Tijuca National Park, Rio de Janeiro City, southeastern Brazil The orange polygon represents the 100% Minimum Convex Polygon (MCP) for all camera traps where red-humped agoutis Dasyprocta leporina individuals were found until August, 2016. The red polygon represents the 95% MCP of localizations of brown-howler monkeys Alouatta guariba until February, 2017.
The first group of howler monkeys (2 males and 2 females) was translocated from wildlife screening centers and private breeders of Rio de Janeiro state to an acclimatization pen in TNP in 2015, and released after three weeks. Two animals of this group were captive-born relatives, while the other pair had an uncertain origin before reaching the screening center. A second group, a wild born male and a captive female, was released in 2016. A third release of one captive-born male was conducted in 2017 – a wild-born female would be released together with it, but had to be removed due to health issues. All individuals, except the last released male (which had recurrent anklet-related injuries) received radio transmitters (TGB-315 and TXE-311C, Telenax®, Playa del Carmen, Mexico; Tigrinus®, Timbó, Brazil), but all transmitters of the first group quickly stopped working. Nevertheless, we have monitored the animals 2–3 times a week, with a field effort of 577h until early 2017, including 347h of actual observation of the individuals.
The reintroduced howlers received supplementary feeding on a tree-platform near the release site, but they quickly ceased feeding off the platform. Fieldwork has focused on monitoring their use of space, diet, and potential effect on morning bird chorus and on dung beetle community, to assess the restoration of ecological interactions.
From the animals released into TNP, two males had to be retrieved and another eventually went missing after his transmitter stopped working. A female of the first group gave birth to a wildborn, which presumably died after four months. After the second group was formed, a female died in March 2017 from unknown causes. The remnant female (the same that had reproduced before) and the newest male bonded in a couple that recently gave birth to another infant. For these last three individuals, home ranges were estimated, and rarefaction curves show they are not expanding horizontally, which suggests these ranges have already been established within TNP (Fig. 2). The survival rates and home range estimates suggest that TNP provides a suitable habitat for a howler population. Our initial low success ratio seems to be related to most animals’ captive origin, and in the next reintroductions we plan to release mainly wild born individuals whenever it is possible.
Restoring ecological interactionsEcological interaction richness increased after the release of agoutis in the park. Immediately after release, the animals fed on more common species and took longer to interact with more rare plant species (Genes et al., 2017). In the first 15 months, they were observed using about 23 out of 65 plant species they were expected to interact with (Genes et al., 2017), burying seeds of large seeded species such as Sterculia chicha (Malvaceae) and Joannesia princeps (Euphorbiaceae) that hardly have another disperser in the park. Seeds of the palm Astrocaryum aculeatissimum germinated only in parts of TNP which had agoutis (Zucaratto, 2013). These results have shown that replicating the reintroduction of agoutis can be an important tool to restore seed dispersal in defaunated Neotropical Forests.
In the first 18 months after release, reintroduced howler monkeys consumed leaves, flowers and fruits from at least 21 tree species belonging to at least 12 families, which is comparable to howler's diet richness in other sites (Diaz and Rangel-Negrín, 2015). Moreover, although the howlers had been absent in the park for more than a century, 25 dung beetle species interacted with their feces, and moved seeds of different sizes, likely enhancing secondary seed dispersal of some large seeded (>10mm) trees in the park. As TNP lacked medium and large sized frugivores, those advances in dispersal can have a disproportional effect on forest regeneration.
DiscussionThe refaunation of Tijuca Forest is an ongoing long-term project, with new releases of howlers, ecological interaction experiments and the release of a third species programmed to occur in the next months. Population reestablishment and the restoration of ecological interactions must be measured during a long period, especially for animals with low reproduction rates as howlers. Nevertheless, we observed some important interactions being reestablished after the two reintroductions, thus refaunation seems to be a promising way to restore ecological processes in the Atlantic Forest.
Our experience with refaunation of the Atlantic Forest also showed us how this process is fraught with difficulties. Although those challenges are not usually mentioned in the literature (Fischer and Lindenmayer, 2000), we believe they are very common in other reintroduction initiatives.
First of all, refaunation practitioners must consider that obtaining the licenses from state and federal environmental agencies needed to start and maintain reintroductions may take longer than expected. To make the relevant bureaucracies more agile would be a very important factor to make rewilding projects quicker and more effective.
Second, source populations may be limiting for several species. The agouti is an exception, with a large source population at Campo de Santana, with over 500 animals. For most other species, the numbers in captive stocks listed in Table 1 may seem reassuring, but this impression can be misleading because not all breeders agree in ceding all their animals for reintroduction. Besides, although animals themselves do not respect state boundaries, to bring individuals from other states is not always an option for genetic (outbreeding) and sometimes legal considerations. In the case of the howlers, we were authorized to use only animals captured within Rio de Janeiro state itself, although it seems debatable whether animals from neighboring areas in São Paulo and Minas Gerais states would actually belong to different populations from a genetic point of view. This source limitation slows the process of creating an initial population, but high survival and home range establishment of individuals released so far lead us to see this reintroduction in a positive perspective despite the long time expected to reach success. Source population limitations was also one of the main problems faced by Bernardo (2012) when trying to reintroduce the red-billed curassow to Rio de Janeiro state. Thus, we strongly recommend efforts to build new ex situ breeding facilities, to improve Zoos and wildlife screening centers and to engage them more actively in reintroduction programs.
Third, many unforeseen difficulties arise during the processes of quarantine, release and monitoring of the animals. For the agoutis, this mostly involved limitations of infrastructure in TNP and Rio Zoo, which caused avoidable losses to the stock for reintroduction, including animals killed by domestic dogs in TNP. Similar problems are likely to be faced by other rewilding projects elsewhere in the Atlantic Forest. For the howlers, on the other hand, some of the challenges that lowered our initial success seem related to their origin – most of them were born in captivity and were used to human proximity. As they are charismatic animals, this lead to some issues regarding tourists and visitors. Three out of the first four released animals got themselves into trouble by looking for interactions with humans, which forced us to retrieve two males and to move two other individuals away from people. To mitigate this problem, we launched an online campaign to warn TNP visitors to avoid feeding the howlers and having any direct contact with them. We expect to have higher success rates in the future by using wild born animals that stood a minimal period in captivity. Nevertheless, especially for social animals, procedures to minimize contact and habituation with humans before release seem highly advisable.
As a fourth issue, there is the problem of genetic diversity. We used Campo de Santana as the single population source for agoutis, as their easy capture there allowed us to intensely harvest the population for reintroduction. On the other hand, in natural populations, estimating the maximum take of individuals and capturing such individuals would be harder. IUCN/SSC (2013) points out the need for genetic diversity of reintroduced individuals to avoid endogamic depression, and because we only used one source, the agouti population is under such risks. However, there have been no signs of low genetic variability affecting population growth and health so far. Nevertheless, all captured animals have their samples of their DNA collected, and genetic analyses are due in the next months. One feasible measure to alleviate genetic problems is reintroducing individuals from other urban parks inside Rio de Janeiro, and there are at least two other sources: Palacete Modesto Leal and Bosque da Barra, although they do not have such large populations. For the howlers, we have not made genetic analyses to identify the variability between different sources of animals, but blood samples were collected from all animals for analyses by the Brazilian Primate Center (CPB/ICMBio).
Reintroductions may have unexpected consequences (Lovari et al., 2014). Because there are no native predators of the agouti in TNP, there is a possibility that their numbers may exceed natural levels and cause seedling damage (Bovendorp and Galetti, 2007), but predation by domestic dogs or food availability may regulate the population. Ongoing population monitoring has shown that so far the population has not increased in an alarming rate, neither in absolute numbers nor in spatial extent.
On the positive side, our experience with rewilding the Atlantic Forest shows that initiatives in this direction can be quite inexpensive. To reach viable numbers of agoutis in TNP, there was an annual cost of USD 6300, for both releasing animals in the wild and monitoring the population (Kenup et al., 2017). This is much lower than the cost of other reported reintroductions, which can go up to a million dollars per year (Fischer and Lindenmayer, 2000). However, it should be noted that this small cost stems from an easily sourceable captive population (the urban population at Campo de Santana). We could not yet reliably quantify the cost for a successful reintroduction of howler monkeys – as the achievement of this goal is still uncertain – and tapirs are surely going to cost even more. Still, it seems likely that refaunation, especially as compared to more widely defined rewilding initiatives using exotics, can be a relatively cheap and effective way to restore ecological processes in defaunated Neotropical forests.
Conflicts of interestThe authors declare no conflicts of interest.
We thank the LECP and LECF teams, especially C. Kreischer, R. Sepulvida, L. Figueira and L. Lima for their help in designing the research, in the fieldwork and in data analysis, and C.H. Salvador for discussions. We thank S.B. Moreira and A. Pissinati (INEA) for their partnership in the howlers’ project and ICMBio/Cetas Seropédica and L. and G. Heliodoro for providing monkeys. We thank L. Bento, the people from Laboratório de Biologia de Tripanossomatídeos/FICORUZ and the staff of Fundação Jardim Zoológico do Rio de Janeiro for quarantine, sedating, veterinary screening and animal manipulation in the agouti project. We are indebted to Fundação Parques e Jardins – Prefeitura do Rio de Janeiro, which provided the agoutis. We thank IBAMA and ICMBio for providing the licenses for the project. We thank the TNP team, especially E. Viveiros de Castro, I. Castro Astor, H. Zaluar, M.L. Figueira, and K. von Kossel for helping in many ways during the process. We also thank to all volunteers who helped collecting data in the field. This work has been supported by Fundação Grupo Boticário de Proteção à Natureza (Programa 0010/2014), CNPq (grant numbers: 487092/2012-4 and 308356/2014-4) and FAPERJ (grant number: E-26/010/001645/2014). We thank the two anonymous reviewers for their suggestions that were very important to improve the quality of the manuscript.

