



Fire has long been recognized as an important driver of vegetation patterns, and is of particular importance for biodiversity maintenance in many grassland systems, including the North American tallgrass prairie. Here, it has been successfully used as a conservation and restoration tool. In southern Brazil, grasslands appear to present many similarities to tallgrass prairie in terms of composition and ecology, but the role of fire has been poorly studied and it is not usually used in conservation. Here, we compare plant genera and family composition of tallgrass prairie and South Brazilian grassland sites. We find striking similarities in terms of dominant families, genera, and functional types. The similarities between plant communities suggest similarities in ecological processes and should lead to a re-thinking of conservation strategies in South Brazilian grasslands. Research on the role of fire is needed, and comparative North–South studies on grasslands in the Americas likely will provide important insights for grassland ecology and management.
Fire is one of the important ecological drivers of vegetation patterns globally and has shaped the emergence of the grass-dominated biomes (e.g. Bond and Keeley, 2005). The tallgrass prairie (TGP) ecosystem located in central North America (Figs. 1 and 2), for instance, is a mesic ecosystem that has been confirmed to be “fire-dependent” (Collins and Wallace, 1990). In the absence of this disturbance, the mesic TGP which receives 700–1100mm of precipitation per year, will transition from grassland to woodland in as little as 40 years (Briggs et al., 2002). For conservation purposes, fire thus constitutes an important tool, not only for maintenance of typical native tallgrass prairie plant communities, but also other conservation components of these ecosystems, such as birds (Fuhlendorf et al., 2006). However, fire is not the only important ecological process shaping prairie communities. TGP plant species diversity is also influenced by the interaction between fire and grazing. Fire singularly in application or in complete exclusion decreases plant species richness, but when combined with grazing the abundances of the dominant grasses are reduced, allowing forbs to increase in abundance (Collins et al., 1998; Coppedge et al., 1998; Fuhlendorf et al., 2006).
 and the South Brazilian Campos grasslands (CSG) in North and South America. We also indicate the location of the Rio de la Plata Grasslands (RPG; see text for details) and study locations from which botanical data was collected. Overlapping points were removed for visibility.")
Location of the tallgrass prairie (TGP) and the South Brazilian Campos grasslands (CSG) in North and South America. We also indicate the location of the Rio de la Plata Grasslands (RPG; see text for details) and study locations from which botanical data was collected. Overlapping points were removed for visibility.

Tallgrass prairie and South Brazilian grassland landscapes. Top: C4 grassland in Oklahoma, USA, in April 2016. The tallgrass in the background is unburned and the green low structure in the foreground is from a prescribed fire ∼35 days prior. Bottom: C4 grassland and Araucaria angustifolia in Parque Estadual do Tainhas in Rio Grande do Sul, Brazil, in early November 2016. The tallgrass in the foreground is unburned and the green low structure in the background is a burn from late August 2016. Note that in both landscapes there is no fence between burned and unburned areas and the preference of cattle grazing in the burned area indicating an interaction between fire and grazing.
Unfortunately, less than 2% of native TGP remains in North America, making TGP a “critically endangered ecosystem type” (Samson and Knopf, 1994; Noss et al., 1995). Given the recognized importance of fire and grazing for maintenance of the TGP, grassland remnants are increasingly placed under a burning regime or a combined burning-grazing regime for conservation purposes (Duchardt et al., 2016). This is especially the case where still large remnants exist, such as in Kansas or Oklahoma, and often in a combination of fire and grazing. Common recommendations are burns every 2–8 years, depending on moisture characteristics with recommendations for more frequent burns in wetter sites (Smith, 2010).
Some 7000km further to the south, the South Brazilian Campos grasslands (SCG) are under subtropical humid climate, and form the dominating vegetation type both in the Brazilian Pampa biome and in the Highlands situated in the southern part of the Atlantic Forest biome (Figs. 1 and 2). The Pampa grasslands are part of the so-called ‘Pastizales del Rio de la Plata’, the extended temperate grassland region that spans over southern Brazil, Uruguay and large parts of northeastern Argentina (Overbeck et al., 2007). Precipitation levels in the SCG are even higher than in the TGP, ranging from 1300 to 2200mm/year (Table 1). Similar to the TGP, these grasslands depend on fire and/or grazing for their maintenance, and when these evolutionary processes are excluded, shrub encroachment and forest expansion take place (Oliveira and Pillar, 2004; Blanco et al., 2014). Thus, the conservation of SCG grasslands might likewise depend on the these disturbance regimes and on their restoration. The use of fire or grazing for conservation, however, is subject to a sometimes polarizing debate (see e.g. Pillar and Vélez-Martin, 2010; Luza et al., 2014; Overbeck et al., 2016). The “forest-bias” in conservation and restoration of the grass biomes is not unique to Brazil but is a common feature in tropical and subtropical regions around the world (e.g. Parr et al., 2014; Veldman et al., 2015). The source of such bias seems to be misunderstandings about the ecology of these systems, in particular the role of disturbances (Veldman et al., 2015). In the case of SCG, this is also evidenced by the use of terms such as ‘steppe’ or ‘savanna’ in Brazil's official vegetation classification, i.e. by problems of ‘taxonomy’ for SCG grasslands.
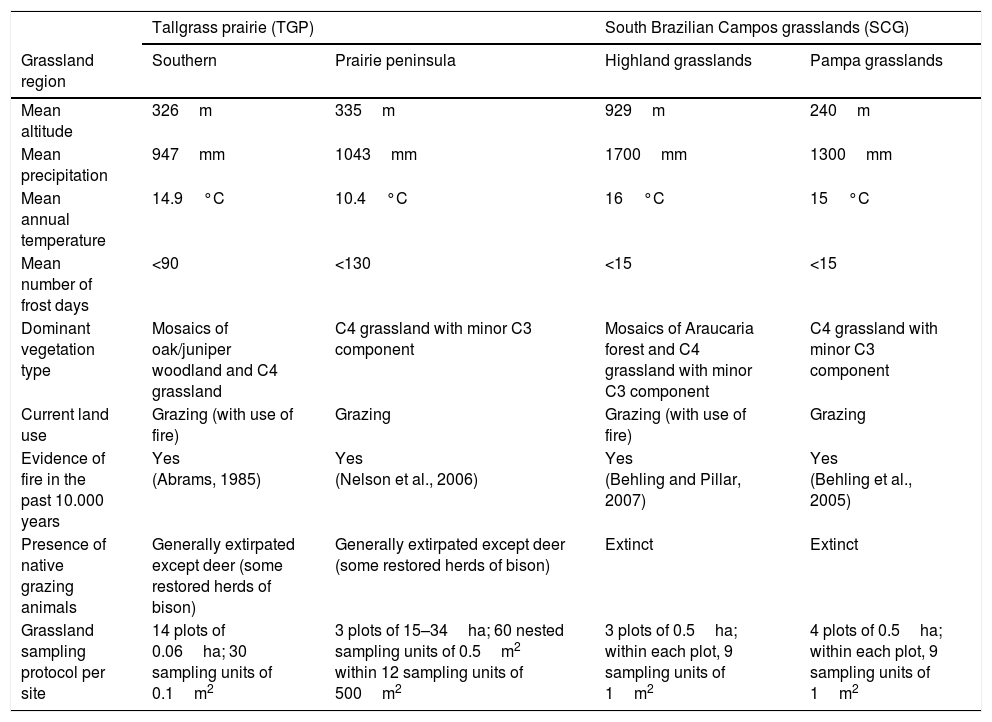
General features of the study regions TGP and SCG.
| Tallgrass prairie (TGP) | South Brazilian Campos grasslands (SCG) | |||
|---|---|---|---|---|
| Grassland region | Southern | Prairie peninsula | Highland grasslands | Pampa grasslands |
| Mean altitude | 326m | 335m | 929m | 240m |
| Mean precipitation | 947mm | 1043mm | 1700mm | 1300mm |
| Mean annual temperature | 14.9°C | 10.4°C | 16°C | 15°C |
| Mean number of frost days | <90 | <130 | <15 | <15 |
| Dominant vegetation type | Mosaics of oak/juniper woodland and C4 grassland | C4 grassland with minor C3 component | Mosaics of Araucaria forest and C4 grassland with minor C3 component | C4 grassland with minor C3 component |
| Current land use | Grazing (with use of fire) | Grazing | Grazing (with use of fire) | Grazing |
| Evidence of fire in the past 10.000 years | Yes (Abrams, 1985) | Yes (Nelson et al., 2006) | Yes (Behling and Pillar, 2007) | Yes (Behling et al., 2005) |
| Presence of native grazing animals | Generally extirpated except deer (some restored herds of bison) | Generally extirpated except deer (some restored herds of bison) | Extinct | Extinct |
| Grassland sampling protocol per site | 14 plots of 0.06ha; 30 sampling units of 0.1m2 | 3 plots of 15–34ha; 60 nested sampling units of 0.5m2 within 12 sampling units of 500m2 | 3 plots of 0.5ha; within each plot, 9 sampling units of 1m2 | 4 plots of 0.5ha; within each plot, 9 sampling units of 1m2 |
The consequences of this lack of understanding of the ecology of these systems and of the forest bias in conservation are readily visible on the landscape – grasslands are much more at risk to transformation into other land uses than forests. Where they are protected, management often is inadequate, as it excludes disturbances (Pillar and Vélez-Martin, 2010). In the South Brazilian Pampa, natural grasslands have suffered a 32.9% decrease in total area from 1975 to 2005, while cover of natural forests increased by 40.7% (Oliveira et al., 2017).
For conservation of grasslands in southern Brazil, a shift toward conservation approaches that include the natural disturbance regimes thus seems necessary, not only to preserve the outstanding grassland biodiversity but also the important ecosystem services provided by them. The application of such approaches should not need to be a “reinventing of the wheel”, but can be based on experiences made in other regions, as long as ecological features and processes are similar. Similarities between SCG and TGP were noted during field excursions in both grasslands within an international collaboration project with participation of researchers and students from the University of Wyoming (UW) and the Universidade Federal of Rio Grande do Sul (UFRGS) in 2016 and 2017. Here, we wish to identify common botanical and ecological features of TGP and SCG that will help us to discuss the ecology of the SCG, to inform potential conservation strategies, and to develop common research needs and conservation challenges and opportunities in both regions.
MethodsWe compare relative cover of plant species, genera and families from SCG and TGP. In SCG, data is from Cambará do Sul, Jaquirana and São Francisco de Paula (Campos de Cima da Serra/Highlands) and from Aceguá, Alegrete and Lavras do Sul (i.e. Pampa). All sites are included in the LTER Campos Sulinos experiment, part of a national network of long-term ecological research studies.1 Study sites are under low-intensity grazing. In the Highland sites, fire is traditionally used for grazing management and at the study sites occurs at intervals of approx. 2–5 years (no detailed fire history data available). In TGP, data were collected in Oklahoma (i.e. southern tallgrass prairie) and Missouri and Iowa (i.e. prairie peninsula). In the Oklahoma site, fire was applied in seasonal treatments with a 2 year interval with no grazing (Weir and Scasta, 2017) and in the Missouri/Iowa site, fire was applied in the late-winter/early-spring with a 3 year interval and seasonal cattle grazing on some plots (patch-burn grazed plots described in Scasta et al., 2016a) and no cattle grazing on other plots. In both regions, our data thus include considerable environmental gradients. General environmental features of the distinct regions are given in Table 1. As we are interested in relative cover of plant species, genera and families, and not in richness, differences in sampling procedures among regions (that may specifically affect the ability to detect rare species) do not constitute a problem. For each site and region, we pooled plot-level data to obtain relative cover per site and region for each species. Plant species were classified according to genera, family, and life form (here in a simple classification into graminoids, forbs, shrubs and trees). Furthermore, grass species were classified according to their photosynthetic carbon metabolism (PCM).
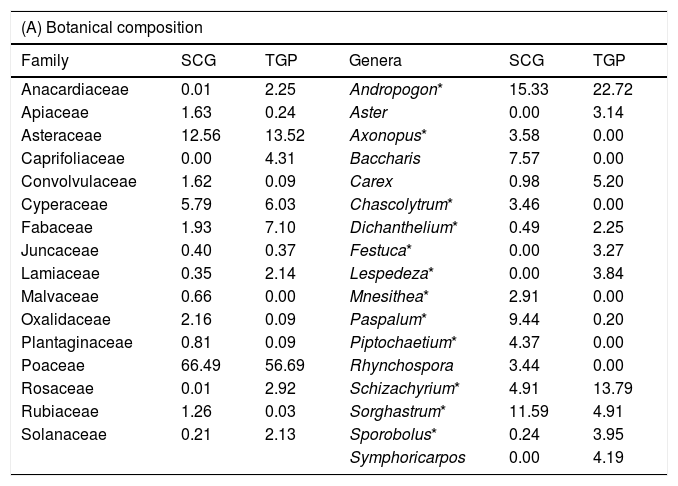
ResultsWe recorded 45 and 61 families from 174 and 190 genera, respectively, in SCG and TGP. In both regions, the families Poaceae, Cyperaceae, and Asteraceae combined contribute to greater than 75% of total vegetation cover (Table 2A). At the genus level, Andropogon showed highest abundance values in both regions with percent cover greater than 15% (Table 2). Graminoids were the dominant life form in both regions, contributing to more than 60% of the total vegetation cover while forbs contributed approximately 20% of the total vegetation cover (Table 2B). Shrubs contributed approximately 8% of the total vegetation cover in both regions. In TGP, we could observe the presence of trees, not present in SCG data, which is likely an artifact of different sampling designs that included a nested sampling design with a final scale of 500 m2 in the TGP specifically to be able to record the tree component (Tables 1 and 2B). Grass portions of the plant communities were dominated by >75% C4 photosynthetic carbon metabolism, and <25% C3 in both regions (Table 2C). The complete list of plant genera from both regions is in the Supplementary Material (Table S1).
Comparison of South Brazilian Campos grassland and tallgrass prairie in terms of botanical and functional composition: (A) Cover (%) of most representative families and genera in each region. Genera marked with * are grasses. (B) Cover (%) and standard deviation (±) of life forms. (C) Cover of grasses with C3 and C4 photosynthetic carbon metabolism (PCM).
| (A) Botanical composition | |||||
|---|---|---|---|---|---|
| Family | SCG | TGP | Genera | SCG | TGP |
| Anacardiaceae | 0.01 | 2.25 | Andropogon* | 15.33 | 22.72 |
| Apiaceae | 1.63 | 0.24 | Aster | 0.00 | 3.14 |
| Asteraceae | 12.56 | 13.52 | Axonopus* | 3.58 | 0.00 |
| Caprifoliaceae | 0.00 | 4.31 | Baccharis | 7.57 | 0.00 |
| Convolvulaceae | 1.62 | 0.09 | Carex | 0.98 | 5.20 |
| Cyperaceae | 5.79 | 6.03 | Chascolytrum* | 3.46 | 0.00 |
| Fabaceae | 1.93 | 7.10 | Dichanthelium* | 0.49 | 2.25 |
| Juncaceae | 0.40 | 0.37 | Festuca* | 0.00 | 3.27 |
| Lamiaceae | 0.35 | 2.14 | Lespedeza* | 0.00 | 3.84 |
| Malvaceae | 0.66 | 0.00 | Mnesithea* | 2.91 | 0.00 |
| Oxalidaceae | 2.16 | 0.09 | Paspalum* | 9.44 | 0.20 |
| Plantaginaceae | 0.81 | 0.09 | Piptochaetium* | 4.37 | 0.00 |
| Poaceae | 66.49 | 56.69 | Rhynchospora | 3.44 | 0.00 |
| Rosaceae | 0.01 | 2.92 | Schizachyrium* | 4.91 | 13.79 |
| Rubiaceae | 1.26 | 0.03 | Sorghastrum* | 11.59 | 4.91 |
| Solanaceae | 0.21 | 2.13 | Sporobolus* | 0.24 | 3.95 |
| Symphoricarpos | 0.00 | 4.19 | |||
| (B) Life form | SCG | TGP |
|---|---|---|
| Graminoid | 72.3±1.8 | 62.9±3.7 |
| Forb | 19.8±1.8 | 26.2±0.3 |
| Shrub | 8.0±1.3 | 7.7±1.1 |
| Tree | 0.0 | 3.2±0.4 |
| (C) PCM (grasses) | SCG | TGP |
|---|---|---|
| C3 | 15.5±1.6 | 22.6±0.9 |
| C4 | 84.5±2.6 | 77.4±4.5 |
The dominant plant family and genera similarity between the two regions was the most conspicuous common feature of both grassland ecosystems during the 2016 hemispherical travel exchange and field excursion (Figs. 2 and 3). Our analyses of the floristic characteristics of the two regions have corroborated these visual similarities. Common and dominant C4 graminoid genera in the South Brazilian Campos grasslands, including Andropogon, Schizachyrium, and Sorghastrum, are also common in the tallgrass prairie of North America. The dominance of these C4 grass genera is indicative of fire (Collins and Wallace, 1990; Uys et al., 2004; Overbeck et al., 2005). The TGP have four dominant species, of which three are within the same genera as dominant species in SCG: Andropogon gerardii, Schizachyrium scoparium, and Sorghastrum nutans (Kelting, 1954; Howe, 1994). Other genera common between TGP and SCG include Aristida, Dichanthelium, Eryngium, and Ruellia (Table S1). Apart from floristics, the two regions are also similar in dominant plant functional types: specifically the mean contribution of C3 and C4 grasses is of the same magnitude, just as that of forb and especially shrub cover. When these botanical similarities are placed in the context of what is known about the fire-dependency of TGP (Collins and Wallace, 1990; Limb et al., 2011; Duchardt et al., 2016; Weir and Scasta, 2017), it suggests that fire as a regulating ecological disturbance could likely be important for conservation in SCG when grazing has been excluded, e.g. in protected areas. Moreover, the integration of grazing with fire to sustain both agricultural livelihoods and achieve complex conservation goals may be a new frontier in SCG where lessons learned in TGP may apply (Noss et al., 1995; Fuhlendorf et al., 2006; Scasta et al., 2016b).
 and burned ∼60 days prior by wildfire (right) at Parque Estadual Do Tainhas in Rio Grande do Sul, Brazil. Bottom: Unburned for more than 2 years (left) and burned ∼90 days prior with prescribed fire (right) at the Grand River Grasslands in Iowa, USA.")
Contrast between burned and unburned grassland. Top: Unburned for more than 2 years (left) and burned ∼60 days prior by wildfire (right) at Parque Estadual Do Tainhas in Rio Grande do Sul, Brazil. Bottom: Unburned for more than 2 years (left) and burned ∼90 days prior with prescribed fire (right) at the Grand River Grasslands in Iowa, USA.
In SCG, we have differences between Pampa grasslands and Highland grasslands regarding dominant grasses: in the Pampa, lower-growing rhizomatous and stoloniferous grasses, such as Paspalum notatum or Axonopus affinis have high dominance, at least under higher grazing pressure (Overbeck et al., 2007). In this region, stocking rates are rather high, which impedes biomass accumulation and leads to dominance of grasses adapted to persist under high levels of grazing, such as the aforementioned grass species. When grazing is excluded the larger caespitose grasses become dominant. The latter are also dominant under frequent burns, such as in the highland region of SCG where Andropogon lateralis, Schizachyrium tenerum and Axonopus pellitus are the most important grasses (Andrade et al., 2016). Here, fire is used because stocking rates in the warm season are rather low, which not only leads to dominance of taller species but also to accumulation of biomass. At sites where fire was excluded for longer periods, the tall Sorghastrum pellitum becomes the most abundant species, at the expense of the smaller grasses and also of the rich forb component (Overbeck et al., 2005). Evidence for the presence of fire in the SCG in the past millennia can be found in peat profiles with charcoal particles, indicating the presence of fire even before human colonization of the region, even though fire events were rather rare (e.g. Behling et al., 2005). One striking difference in our data set seems to be the lack of trees in SCG. However, this reflects the specific sites and sampling procedures from the TGP that were nested at different scales to detect the overstory but the SCG procedures in contrast led to a low change of inclusion of trees in the sampling. For example, at least in the highland grasslands, isolated Araucaria trees are a common feature in the grassland. To include this component in data from vegetation sampling, the use of larger plots or a nested sampling design, as in TGP, would be necessary.
As mentioned earlier, the ubiquitous feature in the Highland region of SCG, the presence of the Araucaria tree, Araucaria angustifolia, is notable from a fire perspective. This gymnosperm tree is the dominant tree in the Mixed Broadleaf Araucaria Forest that can be found on the South Brazilian highland plateau at altitudes above 700m and in mosaics with grassland (Overbeck et al., 2007). However, it does readily colonize grassland where it can occur isolated (Fig. 2) or where it can lead to the formation of small islands of woody vegetation within the grassland matrix (Duarte et al., 2006). Interestingly, the structural characteristics of the Araucaria tree such as its thick bark and its high crown ratio may also be evidence that frequent fires are an ecological process that has structured these mesic tall grasslands for millennia (Behling and Pillar, 2007). However, colonization of grasslands by trees and succession to forest has so far only been addressed from a forest perspective, and not much attention has been paid to the fact that Araucaria trees may also be an element of grasslands. Finally, Andrade et al. (2016) called for more research on the effects of fire and grazing in SCG, in part, because observational and empirical evidence suggests that fire frequency has increased within the last 10,000 years. This increase in fire however cannot be separated from humans because anthropogenic fire has been used as a management tool to remove dry biomass at the end of the winter and likely had been used for hunting purposes by indigenous people as well (Overbeck et al., 2007).
Perspectives and conclusionsTGP and SCG have some regional differences, specially the climate, but share similar botanical, ecological, and social features. Botanically, both are dominated by the same plant families (i.e. Poaceae and Asteraceae), plant life forms, and by C4 grasses. Ecologically, there seem to be strong relationships between disturbances (fire and grazing) and vegetation structure in both regions, as also supported by the available literature (see above). Socially, livestock production is an important (cultural, economic, and social) activity in both regions, and both face the same degradation problems (i.e. conversion of native areas to crops, fragmentation of natural ecosystems, deterioration of native biodiversity, overgrazing, woody plant and exotic plant invasions) (Noss et al., 1995; Overbeck et al., 2007). These similarities lead us to believe that responses of vegetation to disturbance management should be similar. The knowledge made in both regions appears to be relevant to form land-managers’ decisions about which management strategy should be adopted in order to maintain grassland biodiversity and to shape the vegetation according to the system needs which may differ between regions (i.e. forage production, financial feedback) (Scasta et al., 2016b). Additionally, it must be recognized that fire effects are distinct for different plant functional groups and may also vary depending on season of burn (Weir and Scasta, 2017). For the SCG, more detailed studies that evaluate fire effects on specific plant functional groups, and in particular those important for grazing, are missing.
In both regions, there is a demand for not only local but regional-scale studies and global comparisons in order to provide a more general understanding about fire and grazing effects on plant species/community (Weir and Scasta, 2017) and the conservation of grasslands (Scasta et al., 2016b). By analyzing drivers which influence ecosystem dynamics, we can create a comparative framework in order to identify ecological applications that can be applied in different ecosystems sharing common basic features. In this context, studies were carried out comparing disturbance management effects between North America grasslands and South African savannas (e.g. Knapp et al., 2004, 2006; Koerner and Collins, 2014; Kirkman et al., 2014; Forrestel et al., 2015; Smith et al., 2016), and between North America and Australia (e.g. Fujioka et al., 2008). However, there is a lack of studies comparing North and South American grasslands, with the exception of Epstein et al. (2002) which also has minimal focus on South Brazilian grasslands in particular. Fuhlendorf et al. (2006) propose that the fire-grazing interaction should be the basis for grassland conservation in North America. We take this as an indication that perhaps such a fire-grazing interaction framework should be applied in South Brazilian grasslands to enhance conservation of this critical biome. Research on this is clearly needed.
This international cooperation was made possible by a University of Wyoming – Center for Global Studies grant (Travel Exchange to Link Ecology and Production of Analogous Rangelands of the Northern and Southern Hemispheres). Support for the vegetation sampling in North America was provided by Iowa Department of Natural Resources, Iowa Agricultural and Home Economics Experiment Station, and the Iowa State Wildlife Grants program grant #-U-2-R-1 in cooperation with the U.S. Fish and Wildlife Service, Wildlife and Sport Fish Restoration Program (#-U-2-R-1) and the Oklahoma State University Experiment Station. Support for the vegetation sampling in South America was provided by Brazil's Council for Scientific and Technological Development (CNPq; grants 558282/2009-1 and 403750/2012-1 to project coordinator Valério DePatta Pillar). We thank all colleagues and students involved in vegetation sampling in both regions. GEO and IIB acknowledge CNPq productivity grants, and FFF a CAPES scholarship.
The following are the supplementary data to this article:

