





Studies about fish assemblages in Brazilian streams have grown in recent years, however, it remains unclear whether this increase is followed by increments in the diversity of addressed topics and theoretical frameworks adopted by researchers. We performed a systematic search for Brazilian studies on stream fish assemblages recording study region, publication year, objectives, and spatial and temporal scales adopted. The number of studies is unevenly distributed among regions. Most papers describe the general structure of local fish assemblages and their scientific objectives have not varied through time. Studies have been conducted mainly at small temporal and spatial scales, though the latter is increasing over time. We argue for the need of focusing on recently developed ecological theories and frameworks, and expanding the temporal and spatial scales of studies. These changes will improve regional and local conservation policies, and the visibility of aquatic Brazilian research in the global scientific community.
Community ecology seeks to understand spatial and temporal patterns of species diversity and distribution. Throughout the last century, community ecology theory has fully changed from a stable and climax-based concept to a spatially dynamic core determined by local and regional processes (Ricklefs, 1987). Accordingly, the scale of studies on community ecology has considerably shifted, evolving from a small-scale reductionist perspective to a holistic macroscopic view of spatially oriented approaches (Holyoak et al., 2005; Hubbell, 2001). Being informed about the turnover of concepts, ideas, and scales of analyses through time is fundamental for developing new, cutting-edge, theory in community ecology and conservation policies.
Although most of recent ecological paradigm shifts did not come from riverine studies neither by freshwater ecologists, there have been many important developments in the aquatic ecology literature that sought to delve these new ideas within the aquatic realm (Benda et al., 2004; Fausch et al., 2002; Jackson et al., 2001; Grant et al., 2007; Wiens, 2002). Such efforts are important for understanding diversity patterns in fresh waters, particularly in habitats known to contain a large proportion of the biotic diversity and endemisms, such as small headwater streams.
Neotropical fresh waters hold more than 5700 known fish species. About 3200 are present in Brazil (JZ, pers. obs.) and a significant number arguably occur in streams (Castro, 1999). Neotropical streams have been attracting the attention of ichthyologists as a source of new species and sampling efforts in Brazilian streams have therefore increased over the last two decades. A recent meta-analysis identified possible research gaps in Brazilian riverine fish research (Azevedo et al., 2010), though general lines to new stream fish studies are clearly lacking. We develop upon their work and address the following complementary questions: what ecological concepts have been applied in studies of stream fish assemblages and how have they changed over time? How have spatial and temporal study scales changed through time? Answering these questions will guide freshwater ecologists in their future methodological and conceptual choices, providing bases to advance new research lines and also contribute with valuable information to the conservation of stream fishes in Brazil.
We analyzed published research on stream fish assemblages in Brazil to identify (i) the most studied regions, (ii) the main objectives, (iii) the spatial and temporal scales most often analyzed and (iv) their temporal trends over the last two decades. We finally discuss conceptual and methodological aspects that could improve future studies of stream fish assemblages and regional conservation policies. Our research is limited to Brazil and contains most of the information published on stream fish assemblages (except books and papers published in journals not available online), thus representing an up-to-date picture of the Brazilian research on stream fish assemblages.
MethodsWe searched for papers published in scientific journals through the ISI Web of Knowledge and Scielo websites (last assess 02/2012) using the strings “fish*,” “stream,” and “Brazil” in the title, topic, and authors’ addresses, respectively. As we were interested in assemblage-based studies, only papers assessing all species from a sampled location (i.e., fish assemblages) were included, whereas others involving only one or a group of species were not considered. Non-indexed Brazilian journals constitute an important source of stream fish studies, and were therefore included in our analysis when they corresponded to our research criteria (Appendix 1, Table S1).
For each paper, we recorded the year of publication, the Brazilian region where surveys were conducted, the geographic coordinates of sampling sites, the number of sampled streams and sections, and the number of times each site was re-sampled (Table S1). We considered the terms “streams” and “sections” as they were defined in each paper. Overall, streams were distinct, non-contiguous sampling points that were not included in the same channel and separated by confluences; sections corresponded to different sampling points inserted within the same stream.
We recorded the objectives and keywords used by authors in the abstract and introduction of each paper in a presence/absence matrix (Table S1). Studies not testing predictions or patterns from the ecological literature (i.e., studying local richness, diversity, and evenness levels) were considered descriptive and assigned as “Describe the structure of assemblages”. In order to test the variation of objectives over the last two decades, we first applied a non-parametric Multivariate Analysis of Variance (npMANOVA, 999 permutations) using the objective matrix as the response variable and the year of publication as a continuous predictor variable. The Euclidean distance was adopted when computing the dissimilarity matrix. We further analyzed if the diversity of objectives increased through time by applying the Shannon index to the objective matrix and relating it to the publication year with Ordinary Least Square (OLS) models. Finally, we tested if the high frequency of occurrence of the two main objectives could prevent detecting an increase in diversity and variation in objectives through time (Appendix 1).
The temporal scale of each study was defined as the difference between the last and the first sampling year. We used the published geographic coordinates of sampling sites from each paper to estimate the maximum linear distance (i.e., Euclidean distance) between them, representing the spatial scale of each study. When site coordinates were not available (Table S1), we determined the spatial scale directly from published maps by measuring (in cm) the largest distance among the whole set of pair-wise distances between sampling sites and then converting it in kilometers using the scale bar from the published figure.
We analyzed the number of published papers by regions, spatial and temporal scale, and objective category by computing their frequencies of occurrence. We tested for changes in number of streams and spatial and temporal scales over time using Pearson correlations. When necessary, variables were log10-transformed to normalize their frequency distributions.
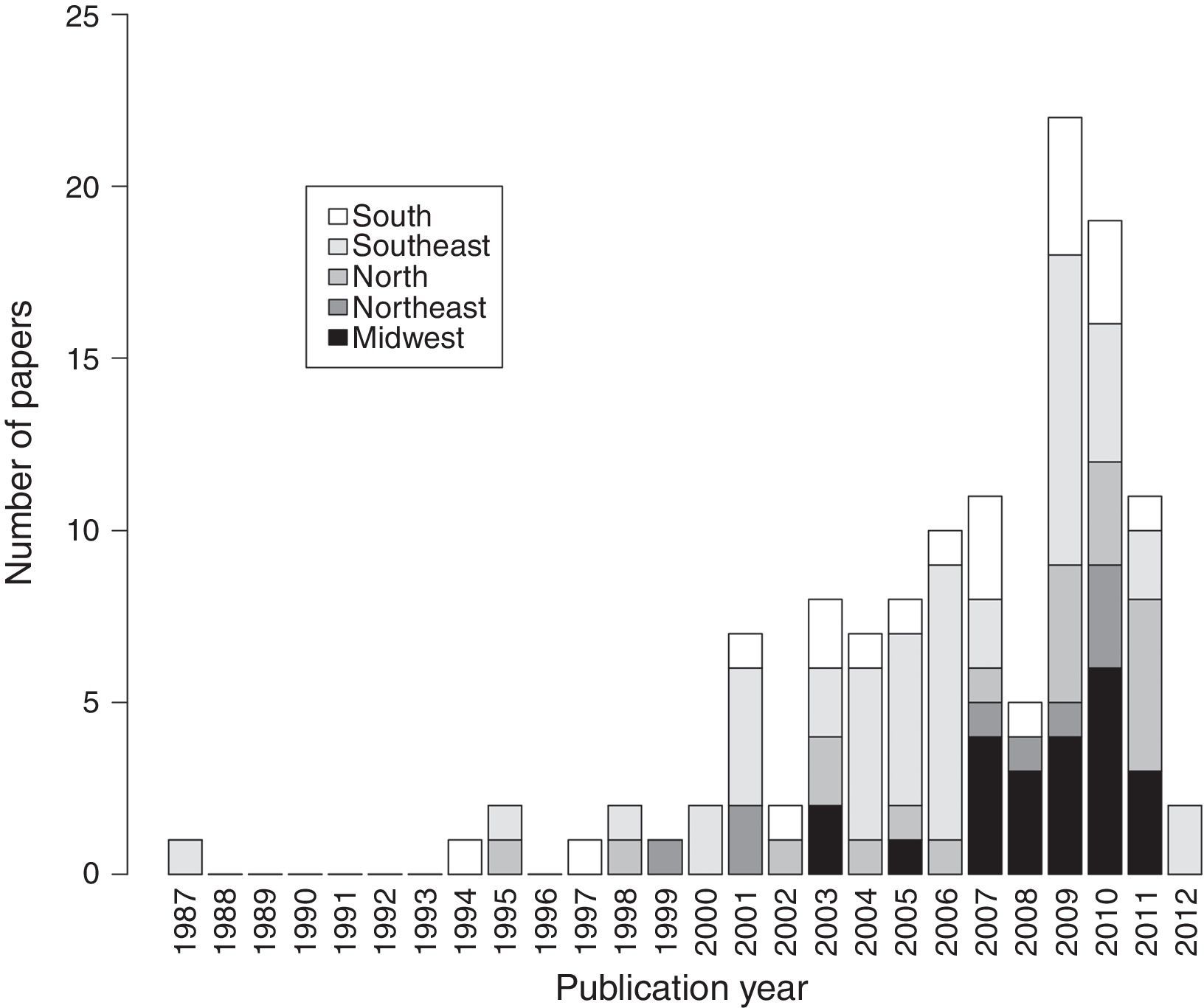
ResultsWe analyzed 124 studies on stream fish assemblages published between 1987 and 2012 (Table S1). The number of papers per year was low but substantially increased after 2000 (Fig. 1). The majority of papers were carried out in the Southeast region (39%), followed by the Midwest (19%), South (17%), North (17%), and Northeast (7%).
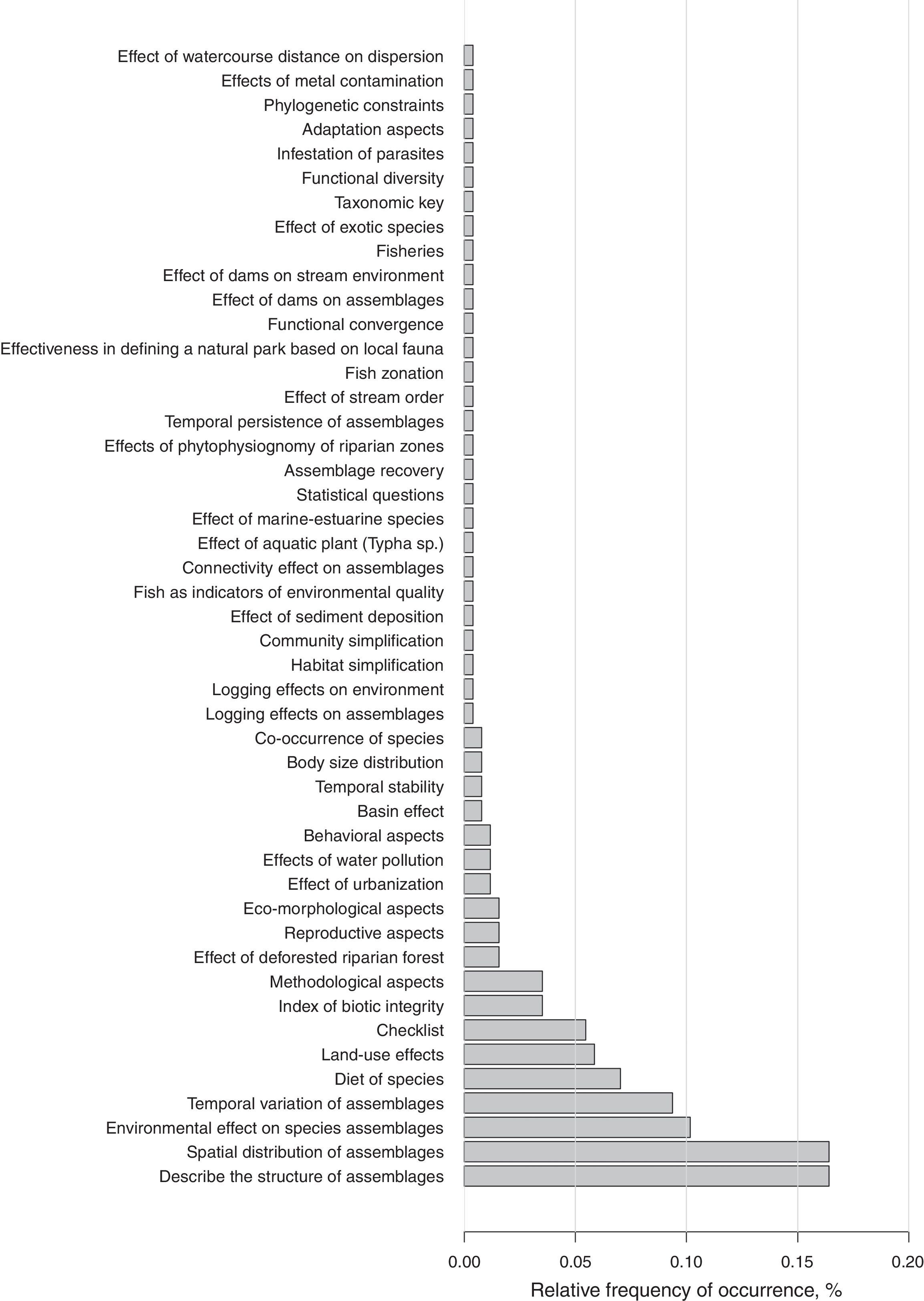
We recorded 48 objectives, though the frequencies of only five of them represent 58% of the total (Fig. 2): “Describe the structure of assemblages” (16%), “Spatial distribution of assemblages” (16%), “Environmental effect on species assemblages” (10%), “Temporal variation of assemblages” (9%), and “Diet of species” (7%). Objectives such as the compilation of species lists, the application of Index of Biotic Integrity, the evaluation of distinct sampling techniques, and the anthropogenic impacts on fish assemblages were recorded at lower frequencies of occurrence (Fig. 2). Despite the large range of objectives, the diversity (OLS, b=0.00, F1,122=0.81, t=0.90, p value=0.37, adjusted-R2=0) and the variation in objectives (npMANOVA, F1,122=1.09, R2=0.009, p=0.36) were not related to year of publication. The lack of relationship for both variables still holds when excluding the two main objectives (Appendix 1).
The mean spatial scale was 62km (sd=108, range=0–685), but the frequency distribution peaked between 0 and 20km and was highly right-skewed (Fig. 3a). The mean temporal scale was one year (sd=1.1, range=0–10) and also right-skewed, with 86% of the studies with one year of sampling or less (Fig. 3b). Moreover, 20% of the studies sampled each site only once, whereas 16%, 16%, and 12% sampled each site four, six, and 12 times, respectively. Most publications studied less than 10 streams (mean=11, sd=23, range=0–200; Fig. 3c). There was an increase in the spatial scale and in the number of sampling sites over time, whereas temporal scale was unrelated to the year of publication (Fig. 3d–f).
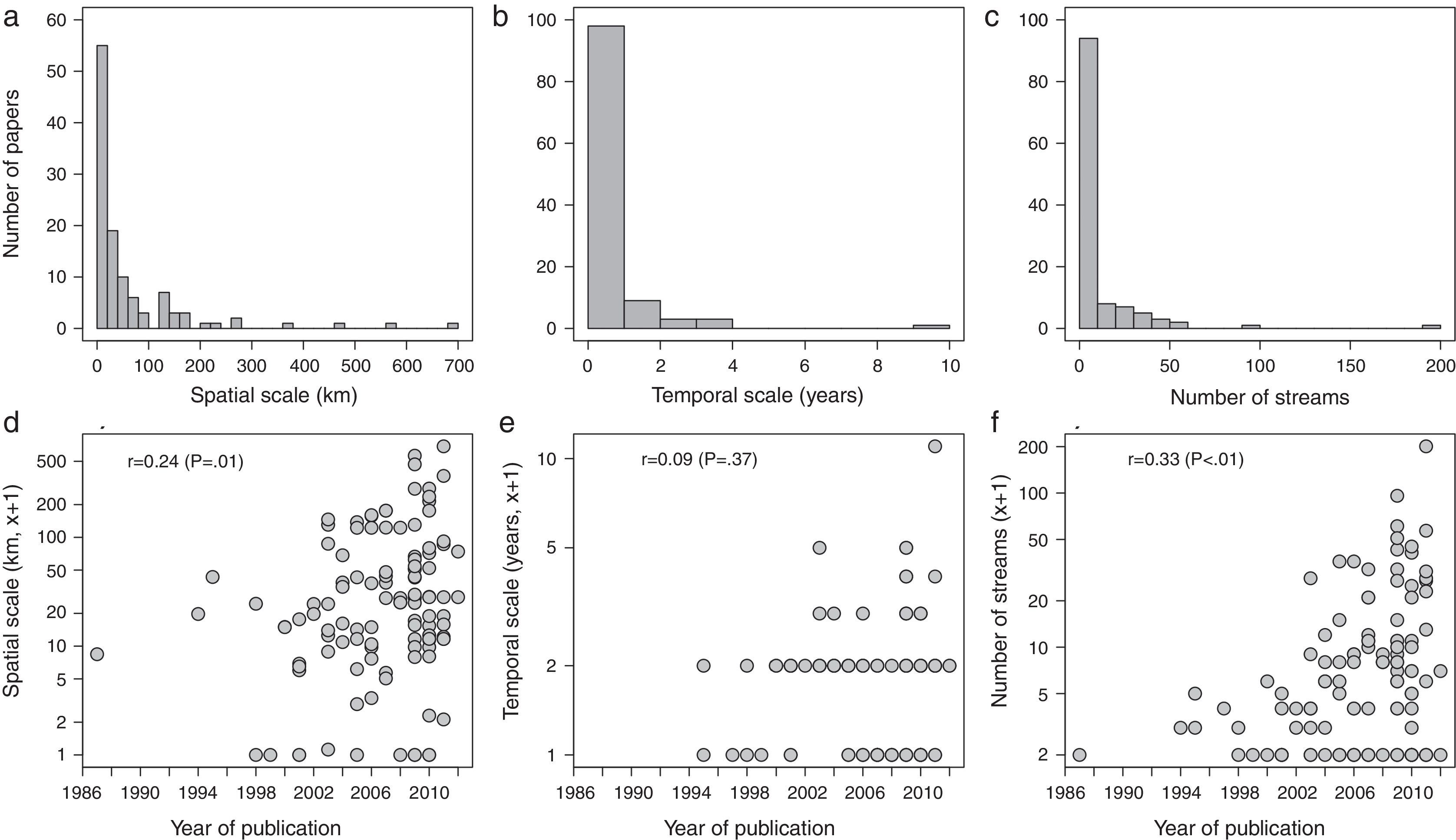
Frequency distribution and temporal changes (respectively) of the spatial (a and d), temporal scales (b and e), and the number of sampled streams (c and f) for Brazilian stream fish assemblage studies between 1986 and 2012. Response variables from plots d–f were log(x+1)-transformed to improve linearity.
Studies on Brazilian stream fish assemblages grew exponentially between 1987 and 2012, with a tipping point around the year 2000, just after the journal Oecologia Brasiliensis published a special issue on stream fish ecology (“Ecologia de peixes de riachos: estado atual e perspectivas,” E.P. Caramaschi, R. Mazzoni, C.R.S.F. Bizerril, P.R. Peres-Neto, eds., v. VI, PPGE-UFRJ, Rio de Janeiro). This influential edition highlighted the importance of streams as a source of fish diversity and has guided much of the subsequent stream research in Brazil. Indeed, there have been key improvements on fish distributional range, biology, diversity, and on fish assemblage seasonal variation, among micro-habitats, variability along the river continuum, and a broad-range of response to local land-use effects (e.g., Objectives/Keywords 4, 7, 9–13, 16–22, 25–26 and references therein, Table S1), all of which constitute the baseline for many of our theoretical expectations and conservation polices for Neotropical aquatic environments. Publications are however unevenly distributed among regions. This pattern may be explained by the greater financial support in wealthy Brazilian states (Azevedo et al., 2010), the higher number of freshwater ecology teams established in Southeast and Midwest regions, and/or the greater number of graduate programs and students in these regions. Indeed, graduate programs and students are important drivers of Brazilian scientific production on biodiversity (Borges, 2008; Nabout et al., 2015). Another important factor is accessibility: streams in Southeast and Midwest regions are located in densely populated regions generally accessible by roads or trails, whereas vast Brazilian areas and their associated aquatic environments remain unevaluated (e.g., in the Amazon).
Studies on Brazilian stream fish assemblages show overall homogeneous objectives and they have not varied over the last 20 years. Most describe general aspects of local assemblages (e.g., diversity, trophic structure, and spatial distribution) and evaluate human impacts on stream fishes (e.g., Dias et al., 2010). Few studies address issues of interest to the international scientific community (e.g., Peres-Neto, 2004) and none test hypotheses related to ecological concepts and theories (e.g., Angermeier and Schlosser, 1989; Benda et al., 2004; Fausch et al., 2002; Muneepeerakul et al., 2008) or tackle the urging issue of climate changes (e.g., Comte and Grenouillet, 2013). Although redundancy in some questions may be beneficial in certain situations (e.g., species’ life history traits at local scales are needed on fundamental grounds and for conservation planning), focusing on a narrow range of topics within a growing scientific community may lead to a conceptual myopia, a partial knowledge gain, and can bring just local static solutions for assemblages facing large-scale threats (Vorosmarty et al., 2010). Overall, these findings may be related to difficulties in recognizing similarities (Fausch et al., 2002; Wiens, 2002) and theoretical limits (Grant et al., 2007) between aquatic and terrestrial ecosystems, the later historically benefiting from stronger theoretical developments. The lack of ecological issues currently debated by the international scientific community, the descriptive nature of most papers, and the absence of significant changes in goals over time could explain, at least in part, the predominance of Brazilian publications on stream fish assemblages in journals with low impact factor (Azevedo et al., 2010).
One approach completely absent from the Brazilian studies is the experimental manipulation of stream reaches. Headwater streams have dimensions (reach length and stream width and depth) allowing for good estimates of species richness, assemblage composition, and population densities, features that can be manipulated in order to test ecological hypotheses and theories in the aquatic realm. The high resilience of fish populations submitted to experimental manipulations (at least in pristine streams; Espirito-Santo et al., 2011) and the low costs of manipulating small streams contribute to the usefulness of stream fishes as models in ecological studies.
Despite the number of papers on stream fish assemblages in Brazil, we show they are mostly performed at small scales relative to the dimension and importance of Neotropical fresh waters. Constraining surveys to small spatial and temporal scales reduces the capability of drawing general ecological conclusions and difficult regional conservation actions. For instance, researchers and managers agree that evaluating the conservation status of most Neotropical fish is difficult due to the limited information on the distributional ranges of species (Polaz et al., 2015). Moreover, better evaluations of the current threats to fish assemblages (e.g., river fragmentation, Perkin and Gido, 2012, effects of climate change, Comte and Grenouillet, 2013) could be done at the landscape scale, thus supporting large-scale policies.
Temporal patterns also represent an essential aspect for understanding freshwater assemblages because some ecological processes usually occur on scales ranging from several years to decades (Hitt and Roberts, 2012). The predominance of short-term studies highlights the lack of regular ecological monitoring in Brazilian streams, which are currently facing multiple anthropogenic threats (Vorosmarty et al., 2010). Moreover, funding agencies often invest in short-term projects (1–3 years), thus favoring short-term ecological studies. Changing the strategy from funding short-term to long-term projects, as well as including streams in existing Long-Term Ecological Programs, could help produce long-term monitoring data, sound ecological results and more comprehensive conservation plans.
Overall, in spite of the predominance of small scales and the gap between the scientific production on stream fish and the ecological theory, Brazilian fish studies seem not to be an outlier. Indeed, much of global freshwater literature is composed of short-term studies and focusing on ecological concepts from the last century (Stendera et al., 2012).
The most urgent need in the Brazilian stream fish ecology is to formulate clear questions incorporating new ecological theories and concepts. Besides recording important descriptive data on local fish fauna, studies tackling new ecological frameworks would enable a deep understanding of the mechanisms driving Neotropical fish assemblages and attain greater impact among the international scientific community. We believe many questions could be tested by using stream fish fauna as a research model (e.g., 38, 40, 43–45, 47, 50–52, and 55 from Sutherland et al., 2013). This would require creativity to circumvent logistic issues and fit theories formulated for, and usually applied to, terrestrial ecosystems to stream reality, thereby producing experimental or analytical innovations. Merging innovative ecological investigations with on-the-ground conservation needs is also desirable and requires an effective communication network between scientists and managers (e.g. Polaz et al., 2015).
To summarize, continuously pursuing new ecological concepts and theories in Brazil is necessary to understand the dynamic and complex processes of Neotropical freshwater systems. The later an overall ecological understanding of aquatic systems is reached, the lower the risk of applying unadapted or cost-ineffective conservation measures. It is equally necessary to increase spatial and temporal scales of studies on Brazilian stream fish assemblages. Extending the spatial coverage of state and federal monitoring programs, the engagement of new/distinct research groups, and possibly the citizen engagement (Dickinson et al., 2012) would facilitate large-scale sampling, inter-ecosystem comparisons, and improvements in under-sampled regions. Even if fish taxonomy is challenging, trait-based approaches could be used to test theoretical predictions, document stream diversity patterns over broad scales (Heino et al., 2013), and to forecast responses to natural and human-related environmental changes. For instance, a trait-based framework would provide knowledge on the functional characteristics of invasive and native species and help to elucidate the roles of biotic factors in the establishment process. Setting permanent national programs of freshwater organisms’ sampling and Biodiversity Monitoring (e.g., those adopted in the USA and France), especially in protected areas, would play a crucial role in generating large-scale data from Brazilian streams. Reaching a larger spatial and temporal vision of freshwater communities of the Neotropics, and the natural and human processes shaping them, would automatically benefit conservation planning, guiding public policies toward the best implementation strategies for preserving aquatic environments, populations and species (e.g. Olden et al., 2010) in a rapidly modifying continent (e.g. Kareiva, 2012). For instance, better integration of ecological relationships from the Theory of Island Biogeography and the metacommunity concept is needed to design effective conservation areas and to adapt them to future distribution shifts as a consequence of climate change. Diversifying the targets of ecological studies, and implementing large-scale sampling designs, together with the uniqueness of Neotropical freshwater fauna, will also promote Brazilian aquatic science to higher scientific levels in the short-term.
Conflicts of interestThe authors declare no conflicts of interest.
We thank CNPq, CAPES, FAPEAM, and INPA (MCTI/PCI/INPA) for funding and U.M. Camargo, B. Hugueny, L. Casatti, J.C. Nabout and one anonymous referee for valuable suggestions on this manuscript. MSD (PhD, Science without Borders program, CNPq/GDE #201167/2012-3; CNPq/PDJ #150784/2015-5) and JZ (#307464/2009-1) received research grants from CNPq. This is paper number #41 of the Igarapés project (www.igarapes.bio.br).
The following are the supplementary data to this article:


