




A research challenge for this century is the integration of highly productive and sustainable landscapes. This issue is crucial for semi-arid regions, where historical land management practices have led to habitat loss and desertification processes. In this study, we evaluated the relative effects of habitat amount (forest cover), landscape heterogeneity (landscape diversity) and spatial arrangement (forest fragmentation and edge density) on bird α and β-diversity in the Caatinga tropical dry forest of northeastern Brazil. We separately assessed the complete bird assemblage and three different ecological groups (forest specialist, habitat generalist and open-area specialist species). Our results indicate that habitat amount is the main positive driver of α and β-diversity of birds in the Caatinga landscapes. However, landscape heterogeneity emerged as an important positive driver for habitat generalist and open-area specialist species. Our results highlight the importance of landscape-scale forest cover and increasing landscape heterogeneity on productive lands as a strategy to balance food production and biodiversity conservation in dry forest regions such as the Caatinga.

Designing sustainable landscapes that conserve biodiversity and produce food, water and energy is a global priority (Williams et al., 2017). Implementing sustainable landscapes requires knowledge of the key drivers of habitat loss and fragmentation in productive landscapes (Laurance et al., 2014). Landscapes embedded in a matrix of anthropogenic activities contain remnants of natural habitats of different sizes and shapes. Thus, landscapes with an intermediate level of disturbance have been shown to have greater spatial heterogeneity promoting biodiversity conservation in productive landscapes (Arroyo- Rodríguez et al., 2020). Therefore, an assessment of species responses is essential to identify optimal scenarios for biodiversity in productive landscapes with higher levels of disturbance (Arroyo- Rodríguez et al., 2020).
Three hypotheses have emerged to explain changes in biodiversity in anthropized landscapes: (i) The habitat amount hypothesis (see Fahrig, 2013) posits that the sample area effect overrides spatial habitat configuration and predicts that the amount of habitat in a local landscape, regardless of configuration, is the main landscape-level driver of species diversity. (ii) The spatial arrangement of habitat patches in human-dominated landscapes suggests that agricultural practices that promote different spatial arrangements of landscapes (e.g., greater structural complexity of landscape form) are often associated with improved habitat conditions for many species and that fragmentation and isolation of natural habitats threaten biodiversity beyond habitat loss (Fletcher et al., 2018). (iii) Habitat heterogeneity suggests that diverse habitats with complex spatial patterns support more species and functions (Martin et al., 2020). However, the evidence for these three main drivers of biodiversity in productive landscapes (hereafter agroscapes) has mainly focused on species richness, but not on other dimensions of biodiversity such as changes in species composition (β-diversity) or species diversity, dominance or the habitat requirements of species.
Ecological traits, which can be specific (e.g., body size, thermoregulatory capacity, or habitat specificity), determine the ability of species to adapt to anthropogenic landscapes (Tonetti et al., 2023). For example, forest specialist species may be more sensitive to landscape-scale habitat availability than habitat generalists (Sekercioglu, 2012). The decline of a particular ecological group can lead to changes in ecosystem functioning (Lu et al., 2024). In contrast, recent research has shown that changes in the composition and configuration of productive landscapes promote changes in alpha (local diversity) and beta diversity (changes in species composition) (Leite et al., 2022). Therefore, it is important to understand how species that perform different ecological functions are affected by habitat loss or increased heterogeneity of land use (Tscharntke et al., 2012).
Decades of research on productive landscapes have led to various land management proposals. For food production and biodiversity conservation in agricultural landscapes, two approaches are often discussed: integrated landscapes that conserve forests and productive areas on the same land, though with lower yields (land sharing), and increasing sustainable production on existing farmland while keeping conservation areas separate (land sparing) (Tscharntke et al., 2012). A theoretical model for biodiversity conservation in productive landscapes proposes a minimum of 40% forest cover, with one large patch (30%) and smaller patches (10%) (Arroyo-Rodriguez et al., 2020). In South America's Caatinga, ideal sustainable landscapes would have 50% natural dry forest and 50% suitable farmland, emphasizing crop diversification and land restoration (Araujo et al., 2020). Recent evidence has highlighted the imperative need to preserve more natural areas in these productive landscapes to sustain essential ecosystem services and mitigate desertification risks from intensive and unsustainable land use (Araujo et al., 2022). However, the direct impact of these models on biodiversity indicators remains unexplored and unproven.
Birds are a diverse and taxonomically well-recognised group, and are important as seed dispersers, pest controllers and pollinators. For this reason, bird communities are used as ecological indicators. (Morante-filho et al., 2015; Sekercioglu, 2012). Forest-dependent bird species in the tropics are sensitive to changes in habitat amount and spatial configuration, but an opposite pattern is observed for non-forest-dependent species (Morante-Filho et al., 2015). Although the positive relationship between bird diversity and landscape heterogeneity is widely accepted, evidence suggests that this relationship varies across geographic regions and biodiversity dimensions (Fluck et al., 2020).
We assessed the relative influence of habitat amount, landscape heterogeneity and spatial arrangement on the diversity of open-area, generalist and forest-dependent bird species using bird field data from the Caatinga dry ecosystems of northeast Brazil. Firstly, we expect that agricultural landscapes with a greater amount of habitat, which are more heterogeneous and less fragmented, will retain a greater α-diversity and β-diversity of birds. Forest species should be more favourably correlated with forest cover, while birds from open areas should show the opposite patterns. Secondly, we expected that the proportion of different land cover types and the distance between samples would be responsible for maintaining β-diversity of birds in the agricultural landscape (Silva et al., 2017).
MethodsStudy areaCaatinga is the largest dryland ecosystem in South America, covering 912,529 km² in the semi-arid hinterland of north-eastern Brazil. Caatinga is an ecological system composed of biodiverse and heterogeneous landscape mosaics containing seasonally dry tropical forests and woodlands (Silva et al., 2017; Araujo et al., 2022). Caatinga has a long history of anthropogenic disturbances, such as the rapid conversion of natural landscapes to commercial agricultural lands (Silva et al., 2017). This study was conducted in Cariris Velhos, the driest subregion of Caatinga in the State of Paraíba (Fig. 1). The climate is hot semi-arid, with rainfall ranging from 350 to 800 mm annually and annual temperature and humidity averages of 25 °C and 65%, respectively (Alvares et al., 2013). About 60% of the rainfall occurs in February, March and April. However, there are large interannual variations. Chronic human disturbance persists in the Caatinga dry environments, with few natural forests and woodlands remaining (Antongiovanni et al., 2020).
Study sites Location of the study area in the Cariri Paraíbano showing land covers and subregion of Caatinga, Brazil, South America, (b) study areas and sampling units (plots) distribution of the radius and points of avifauna sampling in the plots.")
Following the patch landscape approach (McGarigal and Cushman, 2002), we selected 30 plots of 9 ha each as sampling units. These plots were located in landscapes ranging from well-preserved native vegetation to highly degraded areas. (Fig. 1). The land use and land cover were classified using satellite images from Sentinel of 10 meters of resolution (Fig. 1) and field validation was made using GPS. Five types of land cover were identified during the field visits: (i) forest cover, including riverine forest and woodland; (ii) open natural areas, present in the caatinga and rocky outcrops; (iii) cultivated areas, including maize, sorghum and sweet potatoes, peppers, leafy vegetables (lettuce, coriander and spring onions), guava fields; (iv) grazing areas used for grazing; (v) degraded land cover.
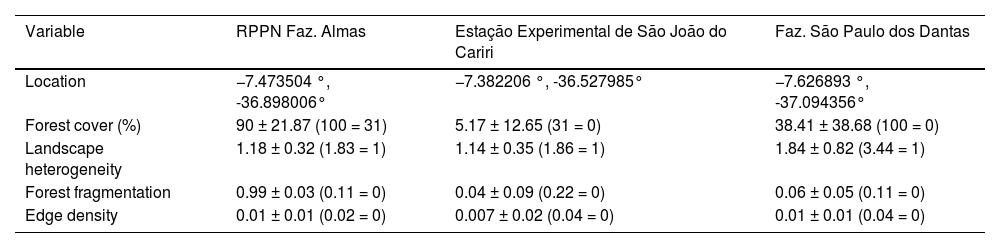
Landscape variablesWe assessed the landscape structure as the quantitative description of the arrangement of elements within a landscape, such as land cover types and fragmentation (Šímová and Gdulová, 2012; Fahrig, 2013) using four measures within each plot: (i) habitat amount, considering the proportion of forest cover and woodland, (ii) landscape heterogeneity, defined in this study as quantitative variation of landscape elements (compositional heterogeneity sensuTonetti et al., 2023), obtained by considering the Shannon land use index (landscape diversity per ha). In addition, we considered two metrics of spatial arrangement, as follows: (iii) forest fragmentation as the degree of fragmentation expressed by the number of forest fragments per hectare, and (iv) edge density as the linear length of forest and woodland measured in km (Table 1). Habitat amount, forest fragmentation, and edge density were calculated using Google Earth Pro, and landscape heterogeneity was calculated using the land use and land cover area and then was estimated in the iNEXT software (Hsieh et al., 2020). These landscape variables have been proposed in landscape ecology studies to identify the most important drivers of biodiversity loss in anthropogenic landscapes. They are widely used, providing high comparative potential (Arroyo-Rodriguez et al., 2020).
Overview of the plots present in the three study areas located in the Cariri region of Paraíba, Brazil.
| Variable | RPPN Faz. Almas | Estação Experimental de São João do Cariri | Faz. São Paulo dos Dantas |
|---|---|---|---|
| Location | −7.473504 °, -36.898006° | −7.382206 °, -36.527985° | −7.626893 °, -37.094356° |
| Forest cover (%) | 90 ± 21.87 (100 = 31) | 5.17 ± 12.65 (31 = 0) | 38.41 ± 38.68 (100 = 0) |
| Landscape heterogeneity | 1.18 ± 0.32 (1.83 = 1) | 1.14 ± 0.35 (1.86 = 1) | 1.84 ± 0.82 (3.44 = 1) |
| Forest fragmentation | 0.99 ± 0.03 (0.11 = 0) | 0.04 ± 0.09 (0.22 = 0) | 0.06 ± 0.05 (0.11 = 0) |
| Edge density | 0.01 ± 0.01 (0.02 = 0) | 0.007 ± 0.02 (0.04 = 0) | 0.01 ± 0.01 (0.04 = 0) |
Bird sampling was conducted between March and June 2019, coinciding with peak Caatinga rainfall and bird breeding season (Araujo et al., 2012). We used four-point counts per plot spaced 200 m apart to prevent double counting, with plots at least 500 m apart (Fig. 1). Each count lasted 10 min, recording birds within a 50 m radius (Araujo et al., 2012), conducted in the early morning hours (between 05:30 and 08:30 h) through visual and auditory identification. We classified all recorded species according to their habitat following Billerman et al. (2020) as open-area species (i.e., those that use native and non-native open areas), generalists (i.e., species found in open and forested habitats like exotic pasture, and agricultural lands and forests), and forest-dependent species (i.e., those that use principally forest habitats) (Table S1).
To assess bird α-diversity at each different habitat use, we used Hill numbers (Jost, 2006): When q = 0 (°D), the index is not sensitive to species abundances, thus representing the total number of species (species richness). When q = 1 (1D), each species is weighted according to its abundance and represents the number of ‘common’ species in the community. When q = 2 (2D), abundances are considered, thus representing the number of ‘dominant’ species in the community (Jost, 2006). Additionally, we performed a sample coverage analysis (Hsieh et al., 2020) to assess whether our sampling was satisfactory (Fig. A.1). These analyses were performed using the ‘iNEXT’ package (Hsieh et al., 2020) from R 4.2.0 (R Core Team, 2022).
Data analysisTo assess the effect of geographical distance on the response variables we use the Moran Index (Moran, 1950). Moran Index allows us to measure how much spatial autocorrelation can be explained by covariance between nearby observations. A non-significant result indicates a spatially random distributional pattern, a significant positive Moran’s I value indicates a tendency toward spatial clustering, while a significant negative Moran’s I value indicates a proclivity toward spatial dispersion. Moran’s was calculated with ape package (Paradis et al., 2019). We found no significant spatial autocorrelation in α diversity (°D: R2 = 0.03; 1D: R2 = 0.01; 2D: R2 = 0.12, P values ≥ 0.1 in all cases).
We analyzed the effect of landscape structure changes as predictor variables (forest cover, landscape heterogeneity, forest fragmentation and edge density) on bird diversity (°D, 1D, 2D) as response variables classified by all species, open-area species, generalists, and forest-dependent species. We used a multi-model approach (Burnham and Anderson, 2004) to calculate the best models that explain the relative importance of each landscape structure metric on each response variable (Giam and Olden, 2016). We calculated Akaike weights (wi), which represent the relative likelihood of a model, and used them to obtain model-averaged parameter estimates (β). We considered models with a difference in qAICc < 2 to have equivalently strong empirical support and similar plausibility (Burnham and Anderson, 2004). The package glmulti was used (Calcagno and de Mazancourt, 2010).
To assess changes in β-diversity at the landscape scale, we used the Bray–Curtis dissimilarity index with results ranging from 0 to 1, 0 indicates that no species are shared between sites and 1 all species are shared (Fig. A.2). We additionally calculated spatial autocorrelation for beta diversity and whether it is explained by habitat amount, spatial arrangement or heterogeneity through generalized dissimilarity modeling (GDM) (Ferrier et al., 2007). The GDM analysis allows us to identify the relative weight of the predictor variables and their independent and combined influence on β-diversity. The gdm package was used (Fitzpatrick et al., 2022). All analyses were made in R 4.2.0 (R Core Team, 2022).
ResultsLandscape structure effects on α-bird diversityThe habitat amount (forest cover) was a more important predictor for α-bird diversity for almost all groups tested, except for open area vegetation specialists where, as expected, it showed a stronger negative effect (Fig. 2). For open vegetation species, landscape heterogeneity has a strong positive effect for species richness (0D) and a slightly less strong but equally important effect for common diversity species (1D) and dominant species (2D) (Fig. 2). As expected, for forest specialist species the largest effect is attributed to the habitat amount available in the landscape for either species richness (0D), common species diversity (1D), or dominant species diversity (2D). However, edge density also showed a strong positive effect on species richness (0D), while landscape heterogeneity and forest fragmentation exhibited negative effects for 1D and 2D.
 complete muti-model including all bird species present in the study area; (b) muti-model including only open vegetation species found only in open habitats; (c) muti-model including only generalist bird species, flexible enough to be used in both open and forest landscapes; (d) muti-model including only forest species. Blue circles indicate positive effects and red represents negative effects. The size of the circles indicates the sum of the Akaike weights (Σwi). The pseudo-R2 shows the percentage of explained deviance for models in the Cariri Paraibano subregion of Caatinga, Brazil. We take as reference the figure from Arasa-Gisbert et al. (2022).")
Effect of the composition and configuration landscape represented in columns on bird diversity in rows. a) complete muti-model including all bird species present in the study area; (b) muti-model including only open vegetation species found only in open habitats; (c) muti-model including only generalist bird species, flexible enough to be used in both open and forest landscapes; (d) muti-model including only forest species. Blue circles indicate positive effects and red represents negative effects. The size of the circles indicates the sum of the Akaike weights (Σwi). The pseudo-R2 shows the percentage of explained deviance for models in the Cariri Paraibano subregion of Caatinga, Brazil. We take as reference the figure from Arasa-Gisbert et al. (2022).
The explained variation of the dissimilarity was 54% by GDM, showing a high variation in species composition between sites. The habitat amount shows the greatest contribution to β-diversity (explained by the increase of the variation in species composition with the increase in forest cover, Fig. 3b). On a lesser but equally important proportion, landscape heterogeneity showed effects on β-diversity (30%), which was more important than geographic distance and the combination of these variables (Fig. 3a).
 of birds for each cover percentage and geographic distance in the Cariri Paraibano subregion of Caatinga, Brazil. The maximum height of each Y axis corresponds to the relative importance of that predictor (in the X axis) to β-diversity. Predictors are shown in order of importance from highest to lowest (left to right, top to bottom). Was used Bray-Curtis dissimilarity index.")
Generalized dissimilarity modeling (GDM) of birds for each cover percentage and geographic distance in the Cariri Paraibano subregion of Caatinga, Brazil. The maximum height of each Y axis corresponds to the relative importance of that predictor (in the X axis) to β-diversity. Predictors are shown in order of importance from highest to lowest (left to right, top to bottom). Was used Bray-Curtis dissimilarity index.
Our results showed that the reduction of available natural habitat drastically affected the total bird assemblage and forest specialist bird species. For open-habitat bird species, on the other hand, we found a positive effect of landscape heterogeneity. Therefore, because bird communities are composed of species with different patterns of response to disturbance, our results reinforce the importance of classifying species assemblages on productive landscapes by ecological traits (Morante-Filho et al., 2015; Vargas-Cárdenas et al., 2022). Our results reinforce the idea that changes in landscape structure and matrix landscape determine the dominance or extinction of functional groups, which in turn influence ecosystem functioning, and are consistent with previous evidence from other regions of the world (Morante-Filho et al., 2015; Lecoq et al., 2021; Vargas-Cárdenas et al., 2022).
Two principal contributions of this study emerge: forest loss in anthropogenic dry forest landscapes is the main driver of bird biodiversity loss, both for total bird communities (in terms of α and β) and for forest specialist and generalist bird species. Furthermore, we detected a positive effect of landscape heterogeneity when open-habitat bird species are considered. This is particularly interesting given that the Caatinga's dry forests have been reduced to their original extent. The remaining forests are highly fragmented at the landscape level and are severely affected by chronic anthropogenic disturbance (Silva et al., 2017; Antongiovanni et al., 2020). Habitat loss remains one of the main threats to bird conservation in the Caatinga dry forest, even though the forest is highly fragmented, and the largest fragments are isolated.
Our results support the habitat amount hypothesis, which suggests that habitat amount is a stronger driver of landscape-scale species richness and diversity than spatial arrangement (Fahrig, 2013). These results are consistent with other tropical forest studies showing that even small fragments of secondary vegetation are important elements for maintaining regional biodiversity (Arroyo-Rodríguez et al., 2020). Promoting connectivity between patches could favor bird conservation in agro-landscapes and subsequent ecosystem services, especially in agricultural landscapes such as the Caatinga (Araujo et al., 2020).
Our results highlight the importance of landscape heterogeneity in mitigating biodiversity loss in human-dominated landscapes (Araujo et al., 2020). In the Caatinga, enhancing vegetation connectivity and crop diversity can support various bird functional groups and ecosystem services. The percentage of land cover, especially forest and pasture + crop, is crucial for β-diversity in tropical agricultural drylands. It is important to note that in our study we used forest cover as the main habitat for forest species, but not for open vegetation species. Although the presence of open-area specialist species is important in the Caatinga (Araujo et al., 2012), our results do not reveal that these species are exclusive to a particular habitat, but they can enhance the landscape in a more generalist way. This result may suggest that open habitat species in the Caatinga may be more resilient to disturbance (Araujo et al., 2020).
Natural habitats in productive landscapes are important for the conservation of α and β-diversity (Fahrig, 2013), for this reason, landscape planning in our study region should prioritize natural forest inclusion and restoration at the landscape level (Araujo et al., 2020; Arroyo-Rodríguez et al., 2020). Nonetheless, the importance of heterogeneous production landscapes highlights the importance of mixed approaches to land management. The importance of spatial heterogeneity and edge density for forest birds in this study underscores the role of landscape elements beyond habitat amount for effective land management (Araujo et al., 2012; Hammill et al., 2018; Martin et al., 2020). For example, edge density has been suggested to have detrimental effects on species that inhabit native vegetation, but the magnitude of the effect depends on the specific species' response (Ries et al., 2004). For this reason, the implementation of management strategies that focus on the ecological characteristics of species in agro-landscapes and the identification of specific elements such as the extent and shape of natural edges can be an important part of management approaches.
According to our findings, landscape sustainability in the Caatinga dry forest requires preserving as many natural habitats as possible, but also promoting heterogeneity on current productive lands. Landscape heterogeneity is linked to the qualitative or quantitative variation of landscape elements, influencing various biotic and abiotic processes such as species diversity in different taxa (Hammill et al., 2018; Martin et al., 2020; Tonetti et al., 2023). Our results confirm that landscape heterogeneity increases β-diversity, especially when forest, pasture and crops are present at the landscape scale, but the current trend of land management in the Caatinga dry forest region promotes accelerated desertification processes (Araujo et al., 2020). Promoting landscapes that integrate both land-based management to conserve and restore natural habitats is a priority. Landscape heterogeneity in productive lands has been proposed as a key management strategy in Caatinga landscapes (Araujo et al., 2020). Therefore, we believe that integrative approaches between separate land management (land sparing) and land integration for production and conservation (land sharing) should be considered in the Caatinga, but in a spatially dependent context (Balmford, 2021).
Conclusions and implications for land managementHabitat amount influences α and β-diversity of birds in tropical dryland agricultural landscapes. While forest habitats boost α-diversity for all groups except open-area birds. Additionally, Landscape heterogeneity supports open-area species, while edge density aids forest specialists alongside habitat amount. Furthermore, besides forest cover, cropland ecosystems enhance bird β-diversity, promoting variation and species diversity in agricultural landscapes. Therefore, we propose general strategies to achieve a balance between food production and biodiversity conservation in Caatinga and other drylands: Conservation and restoration of forest habitats by protecting existing forest areas and restoring degraded areas, and by providing economic and political incentives for the conservation of dry forests. Additional strategies are the conservation of forest remnants adjacent to croplands, variability and crop rotation.
CRediT authorship contribution statementAll authors contributed to the study's conception and design. Data collection in the field was carried out by Mônica da Costa Lima and Helder F.P. de Araujo, and data analysis and interpretation of the results obtained were carried out by Mônica da Costa Lima, Fredy Alvarado and Helder F.P. de Araujo. First version of the manuscript was carried out by Mônica da Costa Lima under supervision of Fredy Alvarado, and Helder F.P. de Araujo. All authors read and approved the final manuscript.
Open research statementData and R code are available in (Lima M. 2023: https://figshare.com/projects/The_importance_of_habitat_amount_and_landscape_heterogeneity_for_bird_diversity_in_tropical_dryland_agrolandscapes/190020
This work was supported by the National Council for Scientific and Technological Development (CNPq; processes n. 441436/2017-0, and n.405298/2021-8). M.C.L. received a scholarship from CAPES Brasil (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior) for a Master's degree within the scope of the Graduate Program in Biodiversity (PPGBIO, UFPB, Campus II, Areia, Paraíba, Brasil; Bolsa CAPES nº 88887.479191/2020 -00). Special thanks to Srs. Delmiro Dantas and José de Arimatéa Braz for collaboration and access to the Farms São Paulo dos Dantas e Almas, respectively.
The following is Supplementary data to this article:
Mônica da Costa-Lima: 0000-0002-2504-3061
Fredy Alvarado: 0000-0002-6724-4064
Helder F.P. de Araujo: 0000-0001-6237-6490
articles

