






Rare and endemic species can be good indicators for monitoring the success of biodiversity management in protected areas. Rarity status can be checked using relatively simple quantitative methods. We used species frequency of occurrence, abundance and biomass to identify rare fish species in a Brazilian Atlantic Forest protected area. We also analyzed the abundance of a vulnerable and endemic species, Isbrueckerichthys epakmos, using general linear models and predictor variables related to instream environmental structures (substrate types, mesohabitats, water velocity and depth). Nine rare species were found, disregarding an exotic specimen. I. epakmos was endemic to PEJU, highly abundant and widely spread species. Its abundance was strongly related to higher proportions of coarse substrate, pools and riffles. The combination of biological information and rarity rates is a useful tool for indicating failures in conservation management, but also for suggesting more effective actions.
In ecological communities, few species are common while most are rare, a consistent pattern across taxa and ecosystems (Magurran, 2004; Magurran and Handerson, 2010a). An intuitive measure of rarity is density, i.e. the number of individuals in a given area (Gaston, 1994). Even though population biomass or the spatial distribution range may equally be employed for expressing rarity, the former is linked to the amount of material and energy flow in food webs (Cohen et al., 2003), while the second is a measure of endemism (Gaston, 1994; Hercos et al., 2013).
Endemic species for example can be good indicators of unique communities, e.g. with many rare taxa (Burlakova et al., 2011). While common species can compensate, in the short term, a decline in ecological functions resulted from a decrease in rare taxa, the loss of additional functions performed by uncommon groups can contribute to further loss of species. It is worth noting that rare species, even those with only one individual in the sample, can sustain vulnerable ecosystem functions, i.e. those that can only be fulfilled by a small group of species with specific traits related to their function (Mouillot et al., 2013). If these keystone species disappear, vulnerable ecosystem processes are also lost, indicating that rare species can influence ecosystem resistance and resilience in the long term (Smith and Knapp, 2003).
Besides the importance for ecosystem functioning, the concept of species rarity is central to biodiversity assessment. For example, species richness estimators assume that the higher the number of species captured in only one sample (uniques) or those represented by only one individual (singletons), the higher total species richness in the community (Chao, 1984; Colwell and Coddington, 1994; Chao et al., 2009). In this sense, all information on unseen species is summarized by the rarity pattern in the samples. Once all species in the sample are represented by at least two individuals (or are presented in two sites) no more species are expected to be found in that community (Magurran, 2010).
When it comes to previously established protected areas – PAs, indicators of rarity can be used to monitor changes in biodiversity patterns over space and time, indicating failures or successes in conservation management (Santiago and Gallo, 2011), whereas for unprotected areas these and other ecological indicators allow detecting priority conservation sites (Metzger and Casatti, 2006).
Unfortunately, basic information about species occurrence and distribution, as well as rarity and endemism rates, are still scarce and unevenly distributed in Brazil (Menezes et al., 2007). Even in currently existing PAs scientific knowledge production is very low, or its content shows theoretical results, with few practical suggestions of management improvement. For the Jurupará State Park – PEJU – the situation is no different.
This Brazilian PA, located in São Paulo State, contributes to biodiversity protection in a large Atlantic Forest continuum. Despite some basic information gathered in the Management Plan,1 current knowledge regarding biotic and abiotic conditions in PEJU is virtually null, according to Santiago and Gallo (2011) evaluation of scientific research applied to the park management. Many reasons contribute to this astounding scenario, particularly bureaucratic obstacles for researches approval and lack of partnerships among researchers and research centers.
The PEJU Management Plan lists 74 fish species distributed in a preserved area that exceeds 26,200 hectares (São Paulo, 2010). Despite activities with high potential for aquatic ecosystems degradation (e.g. housings and damming), there are signs of high levels of endemism in fish fauna. At least four fish species present in the park are listed as vulnerable in the “Fauna threatened with extinction in São Paulo State” (Bressan et al., 2009). This book, published by the State Government in partnership with research centers, used the same criteria and categories proposed by IUCN – International Union for Conservation of Nature. The vulnerable fish species listed in the book and found in PEJU were Isbrueckerichthys duseni (Miranda Ribeiro, 1907) (Siluriformes: Loricariidae), Isbrueckerichthys epakmos Pereira & Oyakawa, 2003 (Siluriformes: Loricariidae), Pseudocorynopoma heterandria Eigenmann, 1914 (Characiformes: Characidae) and Scleromystax prionotos (Nijssen & Isbrücker, 1980) (Siluriformes: Callichthyidae).
In this scenario, we present this study as a contribution to the current knowledge about PEJU fish communities. Rarity status of fish species was assessed using quantitative criteria and the abundance and distribution of the endemic and vulnerable species I. epakmos was analyzed according to instream environmental components.
Materials and methodsSampled stretchesTen sites, 75m long stream stretches, were sampled inside Jurupará State Park and in its buffer zone between June and December 2010 (Fig. 1). This 75m standardized distance allowed us to sample the main mesohabitats (i.e. pool, run and riffles, description in Teresa and Casatti, 2012) and microhabitats (e.g. depths, substrate types, water velocity) within the streams (Oyakawa et al., 2006). Since we aimed to understand the conservation potential of the area, we selected stream stretches in less degraded environments, e.g. with no obvious signs of substrate homogenization or suppression of the riparian vegetation.
Collection of the ichthyofauna and environmental variables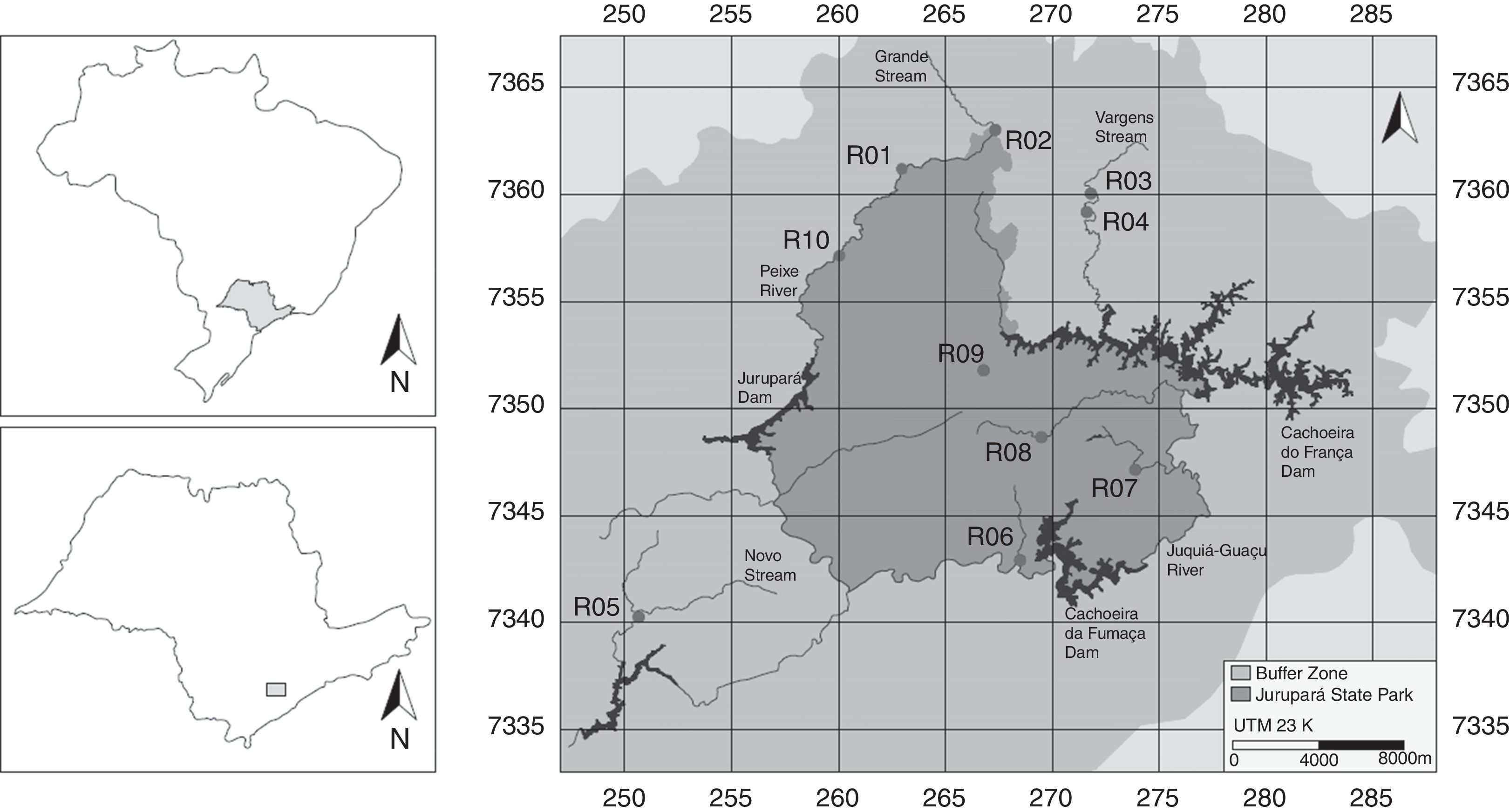
Fish were collected with electrofishing equipment, using a portable generator (Yamaha, model EF2600, 2.3kVA, 60Hz) connected to a voltage transformer. In each site a sweep was made following the downstream-upstream direction, without contention networks at the upper and lower limits. All collected specimens were fixed in formalin for 72h and then preserved in 70% ethanol. Specimen vouchers were deposited in the Fish Collection of Zoology Department in São José do Rio Preto, São Paulo, Brazil.
Three transverse transects were set up at upper, middle and lower limits of each site to guide the quantification of substrate components and channel morphology. Along these transects we visually quantified the percentage of wood debris, pebbles, cobbles and boulders at every one meter (description of all categories in Cruz et al., 2013). The sum of these percentages was used as a variable, named coarse substrate (cs), which was related to bottom substrate stability and microhabitats diversity within the channel (Casatti et al., 2006). Along with substrate components, we also measured channel depth (cm) and water velocity (m/s) and combined the data to calculate a Shannon diversity index for water velocity (H′vel). We added the proportions of the pools and riffles (pr) in each sampled site to create a variable that reflected the combinations of different mesohabitats, capable of housing high species diversity (Teresa and Casatti, 2012).
Data analysisSpecies commonness and rarity were assessed using a modification of the lower quartile criterion of Gaston (1994). The author used the lower quartile of the frequency distribution and the lower quartile of species abundances to classify species as rare. We also considered the lower quartile of species biomass, thus adding another dimension for the classification of rarity. Then, following the recommendation of Magurran (2004) we used the upper quartiles to classify species as common based on frequency of distribution, abundances and biomass.
The frequency of occurrence (Foi) represented the number of sites in which species i was sampled, while species representativeness in terms of abundance was calculated by Ni*=Ni/Foi and representativeness in terms of biomass by Pi*=Pi/Foi (Ni and Pi were total abundance and total biomass, respectively, of species i in all samples). The rationale is that Foi is a proxy for regional representativeness while Ni* and Pi* are proxies for local representativeness in number of individuals and biomass respectively.
Each variable was trisected based on the first and third quartile of the data distribution. Species at or below the first quartile received level 1 (low), species at or above the third quartile received level 3 (high) and the other species received level 2 (medium). Hence, each species received a three-numbers code (for Foi, Ni* and Pi*) which summed defined a score related to its general contribution. For example, a species above the third quartiles for frequency of occurrence, average abundance and biomass was coded as 3-3-3, and its final score was 9. Species with low frequency, low abundance and low biomass received levels 1-1-1 and its final score was 3. These were the maximum and minimum possible scores while other scores varied between these two extremes. Rare species were considered those with a score of five or less. In this case, if a species received code 3 for one criterion it can still classified as rare if received code 1 for the other two. This illustrates for example a large-bodied species (high biomass contribution) that occurs with few individuals and has restricted distribution, or a small-bodied species (low biomass) with high number of individuals but restricted distribution. Common species were those that received scores 8 or 9, so that they must attain high levels in at least two criteria and medium score in the third. The other species were classified as intermediary.
Generalized linear models were used to analyze the abundance of I. epakmos as a function of the percentage of coarse substrate (cs), percentage of pools and riffles (pr) and H′vel (Table 1). These variables were chosen because they are strongly related to the occurrence of Loricariids (Pagotto et al., 2011). We compared models with different combinations of these variables using Akaike second order Information Criterion (AICc) to choose the best model, i.e. the one with the lowest AICc value (Anderson, 2008). Complete method description and additional information are available in Supplementary Material.
General linear models and their respective environmental variables (cs=coarse substrate, pr=pools and riffles, H′vel=diversity of velocity) proposed for analyzing the abundance of Isbrueckerichthys epakmos.
| Model | Environmental variables |
|---|---|
| 1 | Cs |
| 2 | Pr |
| 3 | H′vel |
| 4 | Cs+pr |
| 5 | Cs+H′vel |
| 6 | Pr+H′vel |
| 7 | Cs+Pr+H′vel |
| 8 | Reduced |
Among the 26 sampled species (Table S1), nine scored five or less and thus were considered rare (Fig. S1) – Astyanax ribeirae, Australoheros facetus, Bryconamericus microcephalus, Harttia kronei, Hisonotus sp., Ituglanis proops, Pimelodella transitoria, Neoplecostomus ribeirensis and Neoplecostomus yapo (Table 2). Misgurnus anguillicaudatus was also rare in our samples, but because it is an exotic species it was analyzed separately.
Species grouped according to the levels assigned to each variable (frequency of occurrence, abundance and biomass).
| Characteristics | Levels | Species | Score | Status |
|---|---|---|---|---|
| Restricted distribution, low abundance and biomass | 1-1-1 | 1,3Astyanax ribeirae1Bryconamericus microcephalus1,4Misgurnus anguillicaudatus | 3 | Rare |
| Restricted distribution, low abundance, intermediate biomass | 1-1-2 | 1Pimelodella transitoria | 4 | Rare |
| Restricted distribution, intermediate abundance and biomass | 1-2-2 | 1Australoheros facetus1Hisonotus sp.1Ituglanis proops | 5 | Rare |
| Restricted distribution, intermediate abundance, high biomass | 1-2-3 | Gymnotus sylvius | 6 | Intermediary |
| Intermediate distribution, low abundance and biomass | 2-1-1 | 1,3Neoplecostomus ribeirensis1Neoplecostomus yapo | 4 | Rare |
| Intermediate distribution, low abundance, high biomass | 2-1-3 | Hoplias malabaricus | 6 | Intermediary |
| Intermediate distribution and abundance, low biomass | 2-2-1 | 1Harttia kronei | 5 | Rare |
| Intermediate distribution, abundance and biomass | 2-2-2 | Astyanax sp.2 | 6 | Intermediary |
| Intermediate distribution, high abundance and biomass | 2-3-3 | Astyanax sp.13Deuterodon iguape | 8 | Common |
| High distribution, intermediate abundance, low biomass | 3-2-1 | Characidium lanei | 6 | Intermediary |
| High distribution, intermediate abundance and biomass | 3-2-2 | Hyphessobrycon anisitsi3Geophagus iporangensis | 7 | Intermediary |
| High distribution, intermediate abundance, high biomass | 3-2-3 | Rhamdia quelen | 8 | Common |
| High distribution and abundance, intermediate biomass | 3-3-2 | 2Characidium pterostictum2Phalloceros reisi2Trichomycterus zonatus | 8 | Common |
| High distribution, abundance and biomass | 3-3-3 | 2Gymnotus pantherinus2,3Isbrueckerichthys epakmos | 9 | Common |
Levels: 1 – low, 2 – medium, 3 – high. Score: summed levels. 1Rare species, 2common species and 3endemic species and 4exotic species of the Jurupará State Park.
Eight species were common, i.e. scored eight or nine – Astyanax sp1, Deuterodon iguape, Rhamdia quelen, Characidium pterostictum, Gymnotus pantherinus, Isbrueckerichthys epakmos, Phalloceros reisi and Trichomycterus zonatus (Table 2). I. epakmos is also endemic to the Ribeira de Iguape River basin, which drains PEJU area. Other four endemic species were found: Astyanax ribeirae (categorized as rare), Deuterodon iguape, Geophagus iporangensis and Neoplecostomus ribeirensis (Table S1).
The best model selected to describe I. epakmos abundance included the variables coarse substrate (cs, p<0.001), and pools and riffles (pr, p=0.01). This model was 3.27e32 times better than the reduced model (Table 3).
Model selected by the lowest AICc value to explain the abundance of Isbrueckerichthys epakmos (Iepa). wi=number of times the selected model is more parsimonious than the reduced model, environmental variables: coarse substrate (cs), pools and riffles (pr). p=significance value (95%) of the variable in the model.
| Species | Model | AICc | wi | Variables | Coefficients | p value |
|---|---|---|---|---|---|---|
| Iepa | Cs+pr | 138.56 | 3.27e32 | Cs | 11.41 | 1.24e−10 |
| Pr | 1.18 | 0.01 |
Rare and endemic species are critical components of community structure, in a sense that their loss can result in fauna homogenization (Burlakova et al., 2011), compromising ecosystem functioning (Smith and Knapp, 2003). Several existing proposals to classify species commonness and rarity vary among authors considering different forms of weighing local abundance, spatial range distribution, temporal persistence or habitat specificity (Gaston, 1994; Magurran and Henderson 2003, 2010b; Siqueira et al., 2011; Hercos et al., 2013). Here, we presented a combined classification linking three dimensions of species representativeness at local and regional scales: spatial distribution, abundance and biomass. The cut off level to classify species as rare or common were the first and third quartiles respectively (Gaston, 1994; Magurran and Handerson, 2010a).
Our results show that Jurupará State Park hosts at least nine rare and eight common species. Among the last, I. epakmos showed high frequency of occurrence, abundance and biomass. Its abundance was related to higher proportions of coarse substrate and mesohabitats diversity, i.e. higher proportions of pools and riffles sequences throughout the sampled sites. Habitat diversity within the sampled stretches and preserved riparian vegetation throughout the sites certainly contributed to species occurrence.
Substrate types, from silt to boulders and including wood debris, combined with mesohabitats formed a variety of instream habitats. Among these, shallow riffles, with fast flowing waters and rocky substrate are probably the most suitable habitat combination for I. epakmos. The species has depressed body shape, which decreases water resistance, while the mouth and teeth, similar to a sucking disk, allow individuals to attach themselves to submerged rocks (Covain and Fisch-Muller, 2007). Two other common species, Characidium pterostictum (Characiformes: Crenuchidae) and Trichomycterus zonatus (Siluriformes: Trichomyctheridae), are also rheophilic, i.e. related to riffles, and both are invertivorous active during the daytime (Braga and Gomiero, 2009). Each genus has a strategy for food acquisition, Characidium spp. uses the sit and waits strategy while Trichomycterus spp. actively searches the substrate (Casatti, 2002). These aforementioned species are considered specialists because they are sensitive to habitat degradation (Casatti et al., 2012).
Habitat diversity in PEJU streams also created deeper pools and runs sections, with slower waters and finer sediments. These habitats probably harbored the other two common species, Gymnotus pantherinus (Gymnotiformes: Gymnotidae) and Phalloceros reisi (Cyprinodontiformes: Poeciliidae). Both have less specialized diets and needs in terms of substrate diversity, traits related to generalist species (Casatti et al., 2012). Gymnotus spp. usually settles near the river bank during the day and feeds at night, consuming a wide range of items, from debris to invertebrates. Phalloceros spp. swims near the water surface during the day, feeding on floating items brought by the current (Rocha et al., 2009). Other endemic and common species have generalists habits, as Geophagus iporangensis (Perciformes: Cichlidae) and Deuterodon iguape (Characiformes: Characidae).
Studies have consistently documented how environmental conditions influence species richness and abundance in local scale (Casatti et al., 2006; Teresa and Casatti, 2012; Walsh et al., 2005). Habitat degradation, specially related to substrate simplification, favors generalist species in detriment of specialists (Casatti et al., 2006). Some rare species as Neoplecostomus ribeirensis and N. yapo for example are specialists. The loss of these and other rheophilic species, and the most likely increasing of disturbance tolerant ones, such as omnivores, would probably be the result of fine sediment deposition in runs and riffles. Therefore, maintaining or promoting environmental heterogeneity in small streams must be a priority in PEJU management in order to enhance probability of persistence of specialist and rare species.
Preservation of the riparian zone must be equally important, because it provides wood debris that enhance instream structural complexity, as well as food items (e.g. terrestrial insects) for fish fauna and root mass to stream banks stability (Pusey and Arthington, 2003). Stable banks control sediment loss, consequently preventing substrate homogenization and channel shallowing and widening (Cruz et al., 2013). Therefore, riparian zone affects fish species occurrence directly (Casatti et al., 2012; Teresa and Casatti, 2012) or indirectly, by means of complex interactions with instream conditions (Cruz et al., 2013) so that if degraded, the occurrence of common and endemic species would be in jeopardy.
Endemism can be a good indicator of rare taxa and both groups are essential for communities structure (Burlakova et al., 2011). Our study detected nine rare species in PEJU streams, but certainly there are more. Occurrence and abundance of rare taxa may vary strongly trough space and time, unlike common taxa (Suurkuukka et al., 2012). Thus, increasing sampling effort in terms of geographic range and time periods would probably increase rare species richness and abundance (some species were represented by one single captured specimen, e.g. A. ribeirae, B. microcephalus and P. transitoria). It could also indicate if Misgurnus anguillicaudatus (Cypriniformes: Cobitidae), the exotic specimen, was accidently found in the park or there are signs of a population growth. In this sense, the occurrence of rare taxa can be important in the resistance of communities to new species invasions and colonization (Lyons and Schwartz, 2001).
Naturally, we suggest further studies focusing on the ecological role of rare, common and endemic fish species in PEJU streams. This would help understanding the impact of species loss in the event of habitat degradation. Studies have described how rare taxa can be crucial in nutrient cycling (Marsh et al., 2000) and sustaining vulnerable ecosystem functions (Mouillot et al., 2013), but Gaston and Fuller (2008) also draw attention to common species importance. Despite of what the term suggests, common species are rare in nature. Small declines in their population can cause a huge loss of individuals and biomass, leading to profound changes in communities structure and functioning, as well as ecological services instability.
Our classification approach of species rarity is by no means free of criticism. Samples of tropical invertebrates for example may yield large numbers of singletons or doubletons (Coddington et al., 2009) and Siqueira et al. (2011) noted a similar tendency for tropical stream macrointertebrate communities varying from 50 to 250 taxa. They showed that cutting off only the 25% rarest taxa would disregard several others equally uncommon species and suggest using the inflection point of the species abundance distribution as the boundary between common and rare taxa.
Although this issue deserves further attention, it appears that differences between using the quartile criterion or the inflection point become more acute as the number of species increases, but are less important for communities with few species. Our fish samples yielded 26 species, the half of minimum evaluated by Siqueira et al. (2011) so that few differences would be expected when comparing these two possible forms of classifying species rarity.
Moreover, we combined three dimensions of species representativeness to obtaining a general score of rarity, thus reducing the problem of applying the quartile criteria only top species abundance. For example, species such Gymnotus sylvius or Harttia kronei, that are not bellow the first quartile of numerical abundance were still considered as rare due to their restricted distribution range and low biomass. Finally, an additional advantage of our criteria for rarity is that combining different elements of species representativeness may shed light on different kinds of rarity and on how it is linked with population vulnerability. If a species has lower regional distribution it may indicate a specialist habit. This species would be vulnerable to regional habitat loss from anthropogenic disturbances. Biomass is also important as large-bodied species tend to be less abundant but may have high contribution in transfer of material in community food webs.
In synthesis, our study identified rare, common and endemic species in PEJU streams probably related to instream habitat diversity, which must be preserved. Because riparian zone strongly influences instream conditions, we also reinforce landcover preservation or restoration in degraded streams. Based on our classification, species can be selected as indicators to monitor habitat restoration and biodiversity management in the short, medium and long term. Diversity poor areas may not be resistant and resilient enough, in the future, to withstand environmental changes due to human impacts, exotic species invasions or climate changes. If rare and common species are essential in balancing ecological processes and are not adequately preserved in PAs the entire ecosystem becomes vulnerable. Our classification scheme is easy to be incorporated in management plans of PAs helping identifying species and habitats that deserve special attention in conservation studies. Moreover, evaluating different kinds of rarity (distribution, abundance or biomass) may help to guide more effective actions for preserving different species groups.
Conflicts of interestThe authors declare no conflicts of interest.
We thank CAPES for the MSC scholarship granted to F. A. Teshima and FAPESP for supporting the study “Estudo da integridade física e biótica de riachos das cabeceiras que cruzam a Mata Atlântica Sul de São Paulo” (process 2009/53056-8), of which the present study derived. We also thank Dr. Francisco Langeani for confirming the species, Rinaldo A. Campanhã for allowing our access to PEJU streams and all our colleagues from UFSCar and PUC Sorocaba who helped us during data collection. Finally, we thank the kindness of two unknown reviewers who contributed with valuable comments on preview versions of this paper.
The following are the supplementary data to this article:

The Management Plan is a mandatory technical document that must contain information about biotic and abiotic factors, landscape use and human activities inside the PA and its surroundings. It serves as a guide to natural resources management and governance approaches (Medeiros & Pereira, 2011).

