




In this pioneering study, we evaluated variables that may influence the occupancy and detections (here interpreted as a proxy of intensity or frequency of use) probabilities of cattle and donkeys in a protected area in the Caatinga dry forest biome. No variable influenced the occupancy probability of cattle, which was high and constant at all sampled sites. Conversely, the occupancy probability of donkeys was positively correlated with the distance from the nearest ranch, which might be explained by the fact that donkeys are feral in our study area and thus, avoid areas inhabited by humans. The detection probability of both species was strongly influenced by the type of habitat, with the shrubby arboreal Caatinga being used more intensively by both species than riparian forests. The presence of cattle and donkeys within the protected area may represent a severe threat to the local biotas. This area is currently dominated by secondary successional habitats, resulting from the extensive degradation of its vegetation prior to the establishment of the protected area, and the incursions of these domestic animals may hamper the recuperation of native habitats, as well as jeopardize the survival of its native species, reduce water quality, and erode ecosystem services. We believe that studies of this type are essential and that the data they produce should be considered in the development of the management plans for protected areas to minimize the impacts of these exotic species on the native fauna and flora of Brazilian biomes.
The impacts caused by large domestic herbivores on natural habitats tend to be most evident in arid and semi-arid ecosystems, where water is a scarce resource (Read and Cunningham, 2010; Acebes et al., 2016; Schulz et al., 2016). In these environments, the availability of water has a fundamental influence on the distribution of wild and domestic mammals, especially during the dry season, when this resource becomes even scarcer (Thrash et al., 1995; Smit et al., 2007). A number of studies have shown that cattle (Bos taurus Linnaeus, 1758) and feral donkeys (Equus asinus Linnaeus, 1758) compete with native mammals for space and water sources, displacing them, and impeding their access to these resources (Leeuw et al., 2001; Attum et al., 2009; Acebes et al., 2016).
Native to the arid zones of Africa, the donkey has been successfully established in the arid and semi-arid environments of the New World (Grinder et al., 2006; Vinassa and Nuñez, 2016). This equid was originally introduced into northeastern Brazil during the colonial period (16th Century), with stock being brought primarily from Europe (Salles et al., 2013). Historically, donkeys were used as pack animals and mounts, but have largely been substituted by motor vehicles in recent years. This has led to thousands of individuals being abandoned, and the establishment of a feral breeding population in the Brazilian semi-arid zone (Salles et al., 2013). Cattle were introduced in the Caatinga from the 17th Century onwards, with ranching becoming one of the principal economic activities of the biome, where intense climatic conditions restrict traditional farming practices. These conditions are overcome by the adoption of shifting cultivation, which causes serious damage to the region's natural vegetation and soils (Alves et al., 2009).
Inadequate land management practices, in particular, slash-and-burn clearance of natural vegetation for the establishment of pasture, are associated with the loss of biodiversity (Mamede and Araújo, 2008). Extensive cattle ranching practices also result in the incursion of both cattle and donkeys into forest, where they graze (Schulz et al., 2016). While there has been extensive research on the impact of domestic and feral herbivores on natural environments and the resident fauna around the world (Bagchi et al., 2004; Read and Cunningham, 2010; Peco et al., 2011; Acebes et al., 2016; Hordijk et al., 2019), few data are available on the Caatinga domain (Schulz et al., 2016; Dias et al., 2019), especially in protected areas. Over the past four centuries, ranching has expanded throughout the Brazilian semi-arid zone through the formation of major landholdings. Historically, this activity has included the conversion of native vegetation into pasture, with grazing herds exceeding considerably the support capacity of the environment (Alves et al., 2009). These practices cause severe environmental degradation that is difficult to recuperate, given the characteristics of the Caatinga soils and climate (Santos et al., 2011). In this context, the region's ranchers see the remnants of Caatinga habitat adjacent to their properties as an alternative for the grazing of their herds (Alves et al., 2009). This means that the presence of ranches in the proximity of tracts of forest may have damaging effects on the natural vegetation, may increase the likelihood of natural vegetation suppression, in particular, riparian forests (Santos et al., 2007). While riparian forests are considered important areas for conservation, they are subject to a range of anthropogenic influences and impacts caused by cattle (Costa and Voltolini, 2013). The water and forage availability, as well as amenable temperatures determine the preference of cattle herds for this type of environment (Kauffman and Krueger, 1984; Belsky et al., 1999). However, large herbivores presence in riparian habitats may have negative consequences for the ecosystem structure and the quality of water resources (Strauch et al., 2009).
Given these considerations, we investigated the occupancy probability of cattle and donkeys in relation to the distance from the nearest ranch, the type of habitat, and seasonality in a protected area of the Caatinga in the Brazilian Northeast. We expected to find a negative correlation between the occupancy probability of the cattle and the distance to local ranches, given that these animals are the only domestic stock in the study area. By contrast, we expected to find a positive correlation with distance to local ranches in the case of the donkey, given that these animals are feral in the study area. Here, we considered animals to be feral when they are domestic species but live in the natural environment without any contact with human (Boitani and Ciucci, 1995). We also expected a higher occupancy probability of both species in riparian forest, where water is more available, and a lower general occupancy probability during the dry season, when water is less available within the protected area.
We evaluated the behavioral response of cattle and donkeys to different landscape features by investigating how detection probability, which reflects the frequency (or intensity) of use of an area by a species, varied in relation to habitat variables. Specifically, we expected a negative correlation between the detection probability of the cattle and the distance to local ranches, given that these animals are less likely to frequent areas further from ranches, thereby reducing the detection probability. By contrast, we expected a positive correlation between the detection probability of donkeys and the distance to local ranches, given that this species would be expected to be more frequent in areas distant from ranches, increasing the chances of detection. We also expected both species to be detected more frequently in riparian habitats, due to the greater availability of water in these environments in comparison with the Caatinga forest. Finally, we expected both species to be detected more frequently during the dry season (than in the rainy season), when they range more widely in search of water, increasing the detection probability.
Material and methodsStudy areaThe present study was conducted in the Monumento Natural Grota do Angico (MNGA) with 2183hectare, a strictly protected area of the Caatinga biome, located in the municipalities of Poço Redondo and Canindé do São Francisco, on the right margin of the São Francisco River in the state of Sergipe (Fig. 1), northeastern Brazil. The region's climate is characterized by high temperatures and extremely irregular rains (SEMARH, 2011). During the period of the present study, the rainy season lasted from April to August (data provided by the Sergipe Meteorology Center).
, in the municipalities of Poço Redondo and Canindé de São Francisco, Sergipe, Brazil. The black dots are camera trapping sites; the white triangles are the cattle ranches adjacent to the study area; the São Francisco River is shown in light gray. The geographic distribution of the Caatinga biome in Brazil is shown in the insert (gray area).")
Location of the study area in the Grota do Angico Natural Monument (dark gray), in the municipalities of Poço Redondo and Canindé de São Francisco, Sergipe, Brazil. The black dots are camera trapping sites; the white triangles are the cattle ranches adjacent to the study area; the São Francisco River is shown in light gray. The geographic distribution of the Caatinga biome in Brazil is shown in the insert (gray area).
The predominant phytophysiognomy in the MNGA is deciduous hyperxerophilous forest, with three principal types of habitat: shrubby-arboreal Caatinga, grota (dense forest in ravines), and gallery forest (Silva et al., 2013a), in which the domestic mammals (donkey and cattle) were sampled. In the study area, the shrubby-arboreal Caatinga is at a secondary stage of succession, and its cover is still relatively sparse, and predominantly arboreal. The ravines are associated with intermittent streams with rocky beds, that contain water only during certain parts of the year, making the soil relatively humid, which supports a denser arboreal vegetation. The gallery forest forms in areas where sand is deposited by the São Francisco River, and are influenced by the river flood cycle. The vegetation is predominantly herbaceous-shrubby on sandy soils, with tracts of trees forming corridors in some places. In the present study, we grouped the grota and gallery forests in a single category, named riparian forest.
Data collectionBetween December 2012 and November 2013, we sampled eight sites using digital camera traps with passive infrared sensors, with four traps being set in the shrubby-arboreal Caatinga, and four in the riparian forest. The sites were established at a minimum distance of 500m from one another, and at a maximum distance of 1.5km. The traps were installed along trails at a mean height of 40cm above the ground, and were activated continuously throughout the study period, being checked once a month. To maximize the independence of samples, all photographs of the same species taken by a given trap with a one-hour interval were counted as a single record. Total sampling effort was 2912 trap-days.
Modeling the occupancy and detection probabilities of the species as a function of the covariatesThe occupancy probability (Ψ) is defined as the probability that a sample site i is occupied by a target species (cattle or donkey), while the detection probability (p) is defined as the probability of detecting a target species at sampling site i at time (or moment) t, conditioning the site is occupied (Mackenzie et al., 2002). The detection probability may vary spatially due to habitat characteristics, or temporarily due to seasonal fluctuations in behavior patterns and environmental conditions (Bailey et al., 2004). In this context, here we interpreted the detection probability as the intensity (or frequency) of use (i.e. Cassano et al., 2014; Massara et al., 2018).
At each sampling site, the distance (in km) to the nearest ranch was measured using QGis 2.14 (QGIS Development Team, 2014) and habitat type was identified as either shrubby-arboreal Caatinga or riparian forest. The sampling period was defined as either the rainy or dry season. To determine the influence of these covariates on the species occupancy and detection probabilities, we used a single-season occupancy model (as in Mackenzie et al., 2002) in Program Mark (White and Burnham, 1999). For each camera, we considered whether each species was detected (1) or not (0) during each month that it was deployed, yielding a detection matrix in which the camera was represented for seven dry season and five rainy season months. The detection matrices for the cattle and donkeys were analyzed separately. Finally, we considered the number of days on which each camera was operational at each site during each sampling period to explore for the influence of this covariate on the detection of the probability of the species.
Given the proximity of our sampling sites, we assessed goodness-of-fit (GOF) and evaluated overdispersion (i.e., cˆ), which might be interpreted as a lack of independence among sites, using our most parameterized model and the Pearson's GOF test incorporated in Program Presence (Hines, 2006), which was developed specifically for the single-season occupancy analysis (Mackenzie and Bailey, 2004). We used the Akaike Information Criterion, adjusted for small sample sizes (AICc), to select best models, and considered the covariate(s) of the best ranked models (Δ AICc≤2) to be the most likely determinants of the occupancy and detection probabilities of the target species (Burnham and Anderson, 2002).
We adopted a “step-down” (Lebreton et al., 1992) strategy to build the models. Using fixed structure of a global model (containing all covariates) for Ψ, we built different model structures with only one covariate for p. Based on the best ranked models that contained the most explanatory covariates (Δ AICc≤2) for p, we began to build different model structures with only one covariate for Ψ, fixing the most explanatory covariates for p in a single model. In the final step, we identified the models that provided the best support for the covariates that influenced the occupation and detection probabilities of the target species. We used the maximum likelihood procedures available in the Mark program to estimate the occupancy and detection probabilities for the target species (Mackenzie et al., 2002). Because of model selection uncertainty, we calculated model-averaged estimates of occupancy probability of donkeys (Burnham and Anderson, 2002).
ResultsA total of 440 records of the two-target species were obtained in the MNGA, of which, 142 were of cattle, 109 in the shrubby-arboreal Caatinga, and 33 in the riparian forest. Donkeys were recorded 298 times, with 161 records in the shrubby-arboreal Caatinga, and 137 in the riparian forest. Both species were recorded more frequently during the rainy season than the dry season, with 84 records for cattle and 123 for the donkey.
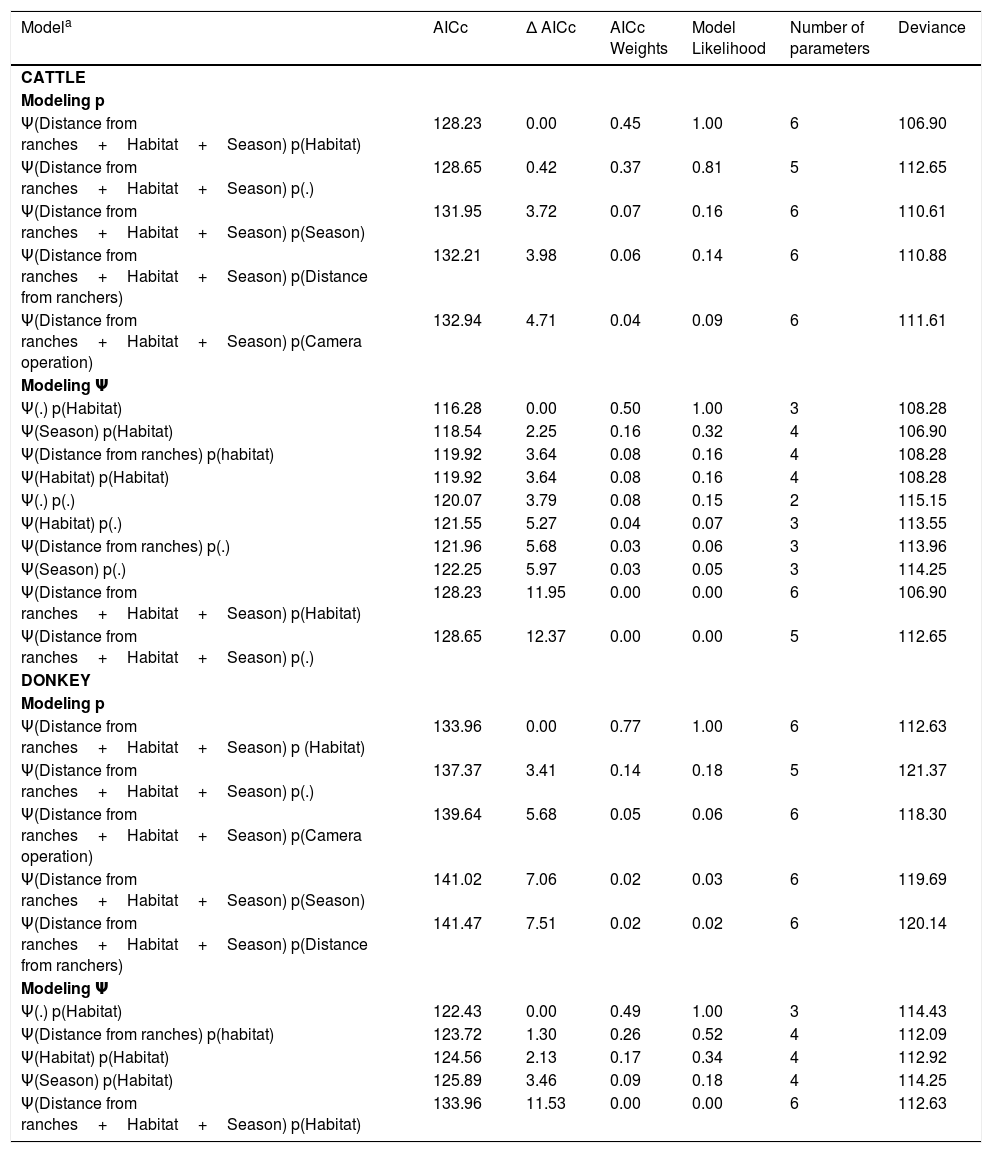
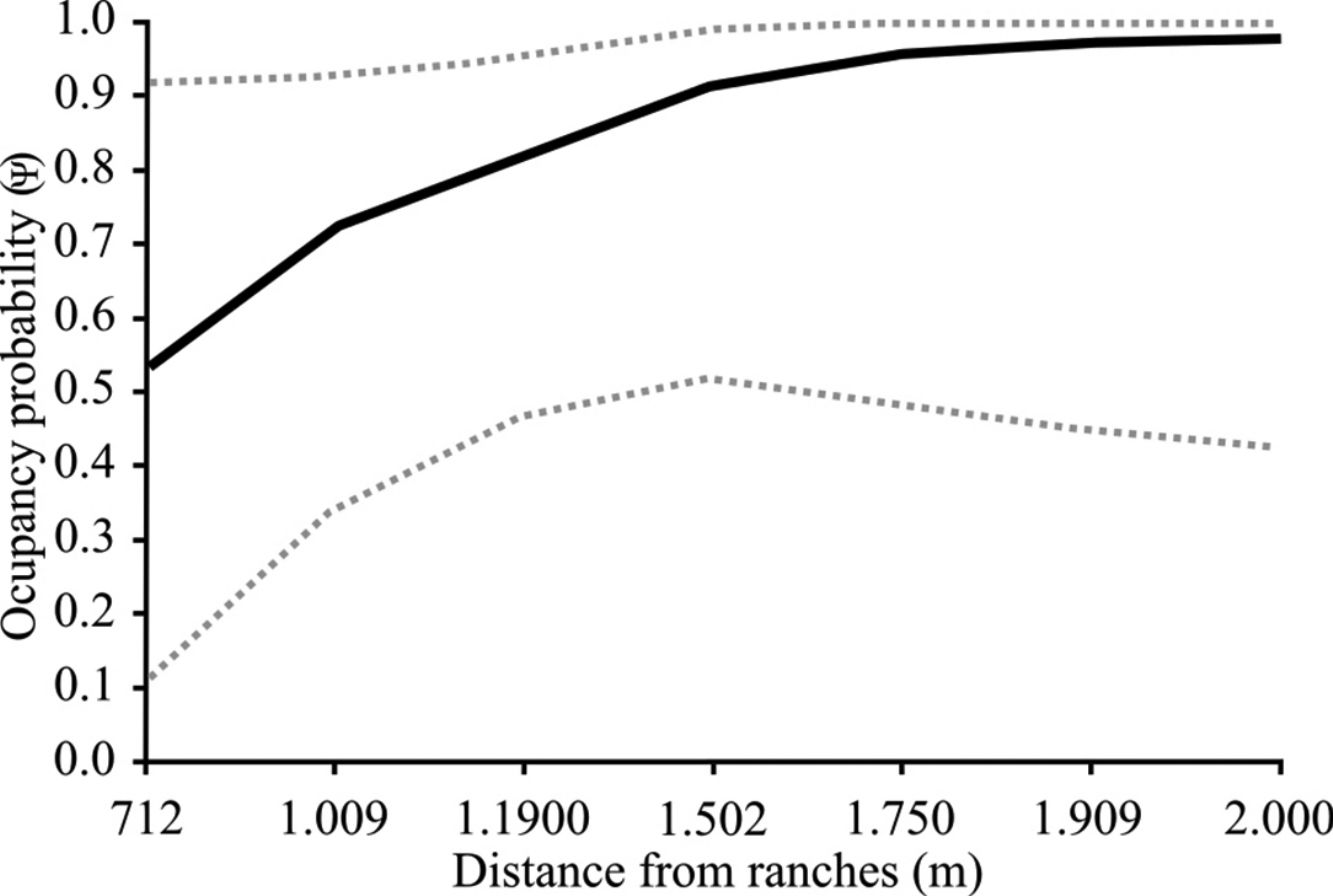
The goodness-of-fit test revealed no evidence of significant over-dispersion (χ2=114.07; p=0.80; cˆ=1.00 for cattle and χ2=199.86; p=0.12; cˆ=1.32 for the donkey), indicating the independence of the study sites. The naïve occupancy estimates (i.e., proportion of sites at which the target species was detected) for cattle was 0.87 and 0.62 for dry and wet seasons, respectively, and the naïve estimates for donkeys was 0.87 and 0.75 for dry and wet seasons, respectively. None of the covariates influenced the occupancy probability of the cattle (Δ AICc>2; Table 1), which was high and constant across all sampling sites (Ψ=0.91, 95% CI=0.36–0.99). We had model selection uncertainty with regards the hypotheses that most influenced the occupancy probability of donkeys (i.e., two models had Δ AICc≤2; Table 1) in the study area. The null (intercept only) model predicted that none of the covariates influenced the occupancy probability of donkeys (Δ AICc=0.00; Table 1), being high and also constant across all sampling sites (Ψ=0.87, 95% CI=0.35–0.99). However, the occupancy probability of donkeys was apparently influenced positively by the distance from the nearest ranch (Δ AICc=1.30; Fig. 2).
Model selection results of the probabilities of occupancy (Ψ) and detection (p) of cattle and asinus in a strictly protected area (Grota do Angico Natural Monument – MNGA) in northeast Brazil sampled in 2012–2013.
| Modela | AICc | Δ AICc | AICc Weights | Model Likelihood | Number of parameters | Deviance |
|---|---|---|---|---|---|---|
| CATTLE | ||||||
| Modeling p | ||||||
| Ψ(Distance from ranches+Habitat+Season) p(Habitat) | 128.23 | 0.00 | 0.45 | 1.00 | 6 | 106.90 |
| Ψ(Distance from ranches+Habitat+Season) p(.) | 128.65 | 0.42 | 0.37 | 0.81 | 5 | 112.65 |
| Ψ(Distance from ranches+Habitat+Season) p(Season) | 131.95 | 3.72 | 0.07 | 0.16 | 6 | 110.61 |
| Ψ(Distance from ranches+Habitat+Season) p(Distance from ranchers) | 132.21 | 3.98 | 0.06 | 0.14 | 6 | 110.88 |
| Ψ(Distance from ranches+Habitat+Season) p(Camera operation) | 132.94 | 4.71 | 0.04 | 0.09 | 6 | 111.61 |
| Modeling Ψ | ||||||
| Ψ(.) p(Habitat) | 116.28 | 0.00 | 0.50 | 1.00 | 3 | 108.28 |
| Ψ(Season) p(Habitat) | 118.54 | 2.25 | 0.16 | 0.32 | 4 | 106.90 |
| Ψ(Distance from ranches) p(habitat) | 119.92 | 3.64 | 0.08 | 0.16 | 4 | 108.28 |
| Ψ(Habitat) p(Habitat) | 119.92 | 3.64 | 0.08 | 0.16 | 4 | 108.28 |
| Ψ(.) p(.) | 120.07 | 3.79 | 0.08 | 0.15 | 2 | 115.15 |
| Ψ(Habitat) p(.) | 121.55 | 5.27 | 0.04 | 0.07 | 3 | 113.55 |
| Ψ(Distance from ranches) p(.) | 121.96 | 5.68 | 0.03 | 0.06 | 3 | 113.96 |
| Ψ(Season) p(.) | 122.25 | 5.97 | 0.03 | 0.05 | 3 | 114.25 |
| Ψ(Distance from ranches+Habitat+Season) p(Habitat) | 128.23 | 11.95 | 0.00 | 0.00 | 6 | 106.90 |
| Ψ(Distance from ranches+Habitat+Season) p(.) | 128.65 | 12.37 | 0.00 | 0.00 | 5 | 112.65 |
| DONKEY | ||||||
| Modeling p | ||||||
| Ψ(Distance from ranches+Habitat+Season) p (Habitat) | 133.96 | 0.00 | 0.77 | 1.00 | 6 | 112.63 |
| Ψ(Distance from ranches+Habitat+Season) p(.) | 137.37 | 3.41 | 0.14 | 0.18 | 5 | 121.37 |
| Ψ(Distance from ranches+Habitat+Season) p(Camera operation) | 139.64 | 5.68 | 0.05 | 0.06 | 6 | 118.30 |
| Ψ(Distance from ranches+Habitat+Season) p(Season) | 141.02 | 7.06 | 0.02 | 0.03 | 6 | 119.69 |
| Ψ(Distance from ranches+Habitat+Season) p(Distance from ranchers) | 141.47 | 7.51 | 0.02 | 0.02 | 6 | 120.14 |
| Modeling Ψ | ||||||
| Ψ(.) p(Habitat) | 122.43 | 0.00 | 0.49 | 1.00 | 3 | 114.43 |
| Ψ(Distance from ranches) p(habitat) | 123.72 | 1.30 | 0.26 | 0.52 | 4 | 112.09 |
| Ψ(Habitat) p(Habitat) | 124.56 | 2.13 | 0.17 | 0.34 | 4 | 112.92 |
| Ψ(Season) p(Habitat) | 125.89 | 3.46 | 0.09 | 0.18 | 4 | 114.25 |
| Ψ(Distance from ranches+Habitat+Season) p(Habitat) | 133.96 | 11.53 | 0.00 | 0.00 | 6 | 112.63 |
 as a function of the distance from the nearest ranch. The estimates are derived from the most parsimonious model that included this covariate, Ψ (distance from nearest ranch).")
The detection probability of both study species was influenced strongly by the type of habitat (Δ AICc<2), with the shrubby-arboreal Caatinga being preferred by both species. Cattle were recorded approximately three times more frequently in the shrubby-arboreal Caatinga than the riparian forest, while donkeys were recorded approximately twice as frequently in this habitat (Fig. 3). None of the other covariates influenced the detection probability of these species in the MNGA (Δ AICc>2; Table 1).
Discussion of the cattle (Bos taurus; A) and donkey (Equus asinus; B) as a function of habitat (riparian forest and arboreal Caatinga). The estimates are derived from the most parsimonious model that included this covariate, p (Habitat).")
The records of cattle and donkeys in the MNGA were 3.7 times more frequent than eight species of medium- and large-bodied wild mammals recorded in the study area during the same study period (see Dias and Bocchiglieri, 2016). None of the covariates influenced the occupancy probability of the cattle, which was high and constant across all sites, indicating that the species is widely distributed, and occurs in all habitats sampled in the MNGA. This pattern was influenced by the management practices of local ranchers, who herd their cattle to the area of the reserve each morning and retrieve them at the end of the afternoon, leaving them to circulate freely within the MNGA during the day. The presence of cattle in protected areas is a widespread phenomenon in the Neotropical region and is a major problem for the conservation of ecosystems, given the impacts caused by these animals on the native vegetation (Piana and Marsden, 2014). Grazing by large herbivores reduces the availability of organic matter for the formation of carbon, which may have a negative effect on the carbon capture of the soil (Schulz et al., 2016). In the context of climate change, the non-sustainable ranching practices that are typical of the Brazilian semi-arid zone are a major concern, especially in protected areas, given that overgrazing is associated with the degradation of the soil and the restriction of a number of ecosystem services (Schulz et al., 2016).
Donkey occupancy probability was higher at sites furthest from ranches, which was expected, given that most donkeys inhabit the area of the MNGA independently of contact with humans. This behavior is typical of feral donkeys, which prefer natural habitats well away from farmland, avoiding conflicts with farmers and possibly because these habitats provide forage (Hamrick et al., 2005; Fuentes-Allende et al., 2016.). Given its ecological plasticity and opportunistic feeding behavior, the donkey can occupy an enormous range of habitats (Reus et al., 2014), feeding on the available resources, either grazing selectively on grass or consuming leaves, bark, and twigs (Fowler De Neira and Johnson, 1985; Grinder et al., 2006). These physiological and behavioral adaptations confer advantages on the donkeys, which allows higher rates of survival than cattle, especially in semi-arid environments (Smith and Pearson, 2005).
These equids tend to spend long periods foraging, and their primary feeding strategy is the ingestion of material with a relatively low nutrient content, allowing them to exploit sources of forage that are underused or even inaccessible to many other herbivores, such as cattle (Mueller et al., 1998; Smith and Pearson, 2005). Given their generalist diet and capacity to exploit fibrous forage, donkeys typically consume more material per unit area than other herbivores and are thus able to survive in relatively small home ranges, similar to those of smaller-bodied species (Seegmiller and Ohmart, 1981). In our study area, feral donkeys present skittish behavior whenever they perceive the approach of humans, braying and then fleeing, although it is not clear why they should avoid humans in the MNGA. Feral donkeys, which have little contact with humans, may develop evasive behavior (McCool et al., 1981) in response to harassment by humans or, in extreme cases, hunting pressure, whether for sport, meat or pest control (Seegmiller and Ohmart, 1981; Reid et al., 1997). As far as we know, however, these activities are absent from the study area. In this case, it would be necessary to investigate the behavior of the feral donkeys of the MNGA to better elucidate the determinants of this behavioral response to the presence of humans. Given these considerations, we believe that the presence of donkeys in the interior of the MNGA is associated with both the avoidance of contact with humans and the availability of forage.
The detection probability of both species was higher in the arboreal Caatinga, contradicting our hypothesis that these animals would be more closely associated with the riparian habitats, due to the availability of water in these environments. This indicates that the presence of these ungulates in this protected area is more closely related to the availability of forage than water resources, which would be related to the fact that the areas surrounding the reserve are highly degraded. Donkeys are adapted to arid regions, and is highly tolerant of thirst (Smith and Pearson, 2005), which enables this species to visit the riparian forest less frequently. In the case of the cattle, this result reflects local ranching practices, with the animals being released to graze in the reserve on a daily basis. The greatest diversity of grasses and other herbaceous plants is found in the shrubby-arboreal Caatinga (Silva et al., 2013b), but while these plants represent an important food resource for cattle in the Caatinga, they are only available during the rainy season (Moreira et al., 2006). As the dry season advances, cacti and the foliage of woody species such as Bauhinia cheilantha and Cenostigma pyramidale become the principal sources of fodder for the animals (Maia and Gurgel, 2013). These plants are very abundant in the shrubby-arboreal Caatinga habitat of the MNGA (Silva et al., 2013b). The seasonality, however, did not affect the detectability of the species. We believe that due to the high degree of degradation of the areas around the MNGA, the accumulated rainfall in the rainy season was not enough for the development of forage plants. In this sense, both cattle and donkeys maintained their frequencies of use throughout the year in the MNGA.
As cattle have been raised extensively in the Caatinga since the 17th Century, many areas of native vegetation are severely degraded by overgrazing (Schulz et al., 2016). The MNGA is typical of this situation, and the ranches and settlements surrounding the protected area are all based on extensive grazing practices. The expansion of cattle ranching into the semi-arid zone of Brazil was based on the conversion of the original vegetation cover by slash-and-burn practices, which causes unsustainable damage on the ecosystem, resulting in the extensive areas of habitat at an advanced stage of degradation observed within the biome in the present day (Alves et al., 2009). As a consequence, reserves of natural habitat that protect native forest are seen by many ranchers as the only potential sources of forage available for their cattle and, in some cases, of water as well (Silva et al., 2013a). The presence of large domestic herbivores in natural areas causes a number of impacts, including the compaction of the soil, the reduction of undergrowth and leaf litter cover, the modification of water quality through the contact of feces and urine with bodies of water, and the propagation of exotic plant species (Santos et al., 2007; Miranda and Muniz, 2009; Costa and Voltolini, 2013; Hordijk et al., 2019).
The presence of cattle and donkeys in the MNGA may exacerbate these impacts, creating a major problem for the conservation of the area, given that much of the local habitat is still undergoing secondary succession, after a long history of deforestation and burn-off within the area prior to the creation of the conservation unit (Silva et al., 2013a). Constant grazing by large ungulates has also been shown to have a negative effect on the species richness and abundance of small mammals (Pedó et al., 2010; Read and Cunningham, 2010), and a recent survey in the MNGA revealed a reduced species richness of small mammals (Freitas et al., 2017). Studies have shown that the suppression of the vegetation by cattle can impede the establishment of communities of small mammals by reducing the availability of feeding resources and refuges, making these animals more vulnerable to predation (Torre et al., 2007; Read and Cunningham, 2010). Cattle and feral donkeys may also affect the communities of larger-bodied wild mammals in the MNGA (Dias and Bocchiglieri, 2016). The spatial overlap between large domestic mammals and wild animals may often by the principal mechanism of exploitation competition, especially in environments where resources are scarce (Acebes et al., 2016). The donkey, for example, is widely distributed in the arid ecosystems of the New World, where the territorial behavior of males may have a negative impact on the use of space by wild mammals, in particular other herbivores (Moehlman, 1998). As generalists foragers, donkeys will also consume virtually any type of plant material (Grinder et al., 2006), including woody plants and cacti, affecting the reproductive parameters of their populations (Malo et al., 2011; Peco et al., 2011).
While the impacts caused by the presence of cattle in natural areas are well known, ranching is an integral component of the region's rural way of life, and an important element in the socioeconomic structure of its communities (Wangchuk, 2002). This emphasizes the need for the understanding of local problems and the identification of alternatives that can guarantee the coexistence of cattle ranching and conservation aims (Lúcio et al., 2014). The extensive ranching practiced traditionally in the region does not contemplate fallow periods to allow for the recuperation of the forage and, as the density of animals generally exceeds the carrying capacity of the habitat, environmental degradation is the standard result (Miranda and Muniz, 2009). Sustainable grazing can only be achieved through the application of management practices that permit the recuperation of the pasture (Melado, 2007). Rational grazing technology may nevertheless be capable of conciliating the needs of the animals with conservation objectives. Productivity can be maintained when ecological grazing practices are compatible with the maintenance of a region's biodiversity (Enri et al., 2017).
In this context, a number of measures can be recommended to minimize the impact of the domestic species on the native biota of the MNGA, while guaranteeing the needs of the ranchers and farmers on the properties adjoining this protected area. The installation of gates and cattle guards on the access roads to the reserve would be one important, low-cost measure that would have immediate results, for example. It would also be important to advise and conscientize local ranchers with regard to the illegal nature of releasing cattle into a strictly protected area. In addition, it is essential that certain measures contained in the MNGA management plan are put into practice, including the permanent and systematic monitoring of all the different zones of the reserve, and the maintenance of its limits. Given the risks of disease transmission, it is also fundamentally important that effective measures are established to monitor and control the introduction of exotic species, with the participation of the local population, to avoid possible conflicts between the conservation of the region's biodiversity and the exploitation of its natural resources.
Although we had an apparently limited number of camera sites, we did not have any lack of fit when constructing our model set (i.e., even our fully parameterized model fits the data). However, we would like to suggest that future studies increase the minimum distance (e.g., 1-2 km) between camera sites to encompass all studied area, but still using the same number of camera sites. Finally, we would like to emphasize the importance of the pioneering approach adopted in the present study for the assessment of the occupancy probability of large domestic and feral herbivores in a protected area in northeastern Brazil. We hope that a similar approach can be applied not only to other Caatinga sites, but in similar areas in other Brazilian biomes where natural habitats are being converted into an anthropogenic matrix. In this context, our findings should provide an important database for future studies, considering that the understanding of the occupation of natural areas by potentially harmful species is fundamentally important for the administration of these areas, and should be considered during the decision-making process for the development of effective management plans for protected areas.
We are grateful to Cenap/ICMBio and the Neotropical Grassland Conservancy for Research Equipment Program. We would also like to thank SEMARH/SE (license # 032.000-01117/2012-4) for logistic support (infrastructure and material), and the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) provided Student Grant Program to DMD. We are also grateful to sr. Didi and James Cardoso for assistance in field activities and the anonymous reviewers for valuable suggestions for improving the manuscript.

