
South America is home to some of the largest, most diverse grasslands on the planet, providing critical habitat for numerous plants and animals and vital resources for millions of people. Yet, at the dawn of the 21st Century, South America's grasslands are experiencing rapid, widespread and unprecedented impacts from activities such as pesticide use, drainage, urbanization, and conversion to industrial monocultures. In combination with the pervasive effects of global climate change, these impacts have the potential to ripple across grassland food webs in ways we still poorly understand. How do these impacts affect the behavior, ecology and population demographics of the numerous migratory birds that call these grasslands home? Given the historical lack of research on grassland birds across the New World, we know very little about how to best conserve them, much less which grassland migratory birds are currently impacted or likely will be in the near future. We argue that, by applying recent theoretical and technological advances in the field of movement ecology, rapid advances can be achieved in understanding grassland migratory birds in South America. We outline the current state of knowledge about grassland birds that migrate within South America, the main threats to their survival, and how the emerging field of movement ecology can contribute to understanding how to conserve and manage their populations. Only with information on the full annual cycle of these birds will we be able to formulate effective conservation and management plans to ensure the long-term viability of these unique species.
South America is home to a highly diverse array of grasslands, from relatively small grassland ‘islands’ located within larger forested landscapes, to high elevation Andean Puna and Páramo, to extensive lowland ‘seas’ of grass such as the south-temperate Pampas and tropical Llanos (reviewed by Vickery et al., 1999; Azpiroz et al., 2012). Given South America's status as the planet's cradle of bird diversity, it is not surprising that the continent's broad spectrum of tropical and temperate grasslands play host to a taxonomically, behaviorally and ecologically diverse set of bird communities, which have been adapting to these open ecosystems for millennia (Ribeiro et al., 2016).
Yet, with few exceptions, South America's grassland birds have been poorly studied. Although the last decade has seen an increase in research on grassland bird communities (e.g., Azpiroz and Blake, 2009; Di Giacomo et al., 2010; Medan et al., 2011; Isacch and Cardoni, 2011; Codesido et al., 2013; Dias et al., 2013; Dotta et al., 2015; Fontana et al., 2016), we still know very little about such basic aspects of their biology as life history strategies, winter habitat use, movements and evolutionary history (e.g., Jahn et al., 2009; Azpiroz et al., 2012), in large measure due to a lack of long-term, standardized schemes to monitor their populations. In most cases, we can only guess at the drivers of their population dynamics or of the effect on their populations of recent, widespread and rapid changes to their native habitats. Compared to many resident species, migratory birds in the Neotropics tend to have large wintering ranges (which has been shown in European breeding species to be negatively related to population declines, Gilroy et al., 2016) and lower habitat specificity, such that they have generally been of lower conservation concern than residents (Stotz et al., 1996). However, some grassland migratory birds are year-long habitat specialists, especially those occupying native grasslands and marshes, such as Sporophila seedeaters (Stotz et al., 1996; Silva, 1999).
We also know very little about the role of South American migratory birds within their ecosystems (but see Bravo et al., 2015). Migratory organisms, including birds, often provide key physical links between habitats and ecosystems, for example transporting genes and propagules (e.g., Viana et al., 2016a, reviewed by Viana et al., 2016b), which can influence gene flow and lead to the establishment of new species (Bauer and Hoye, 2014). Because of the large number of birds involved (i.e., millions of individuals) and distances they travel (i.e., hundreds to thousands of kilometers), many grassland migratory bird species in South America likely provide vital ecosystem services to the grassland system(s) they inhabit (e.g., Swainson's Hawks, Buteo swainsoni, feeding on grasshoppers while overwintering in south-temperate grasslands; Sarasola and Negro, 2005), as has been shown in other habitats (Şekercioğlu et al., 2016). Given their likely important role in maintaining intact, functioning grassland ecosystems, there is an intrinsic merit in understanding and conserving migratory birds in these systems. In turn, if the movement of grassland birds affects the functionality of grassland ecosystems, then understanding their movements provides vital important information on the best conservation practices for these ecosystems (reviewed by Mitchell et al., 2013).
There is growing recognition that understanding how organisms move in space and time is imperative to a full appreciation of their evolution, behavior and ecology, as well as an ability to identify their vulnerabilities on a rapidly changing planet (Marra et al., 2015; Wilcove and Wikelski, 2008). This new paradigm, combined with rapid technological and methodological advances in remote information gathering and analysis, is leading to rapid growth of movement ecology research (Nathan et al., 2008). In spite of the paucity of information on the movements of South America's grassland migratory birds, the application of existing movement ecology theories and tools for data collection and analysis can remedy this important gap in our knowledge of this group of birds and how to conserve them.
Our first objective here is to provide a brief overview of South America's grasslands and their migratory birds, outlining known or potential threats to their populations. Second, we describe how the field of movement ecology, including its theoretical framework and analytical methods can improve our understanding of South America's grassland migratory birds, and consequently, the information necessary to develop effective conservation and management planning for their populations. Although research at the species level is important for promoting grassland bird conservation (e.g., Azpiroz and Blake, 2009; Dotta et al., 2015), we focus here on movement ecology research at the individual and population levels, since an individual's behavior ultimately impacts processes at the population level. For example, populations can be negatively impacted when individual migrants do not correctly time their arrival on breeding grounds to match changes in food resource availability (Both et al., 2006).
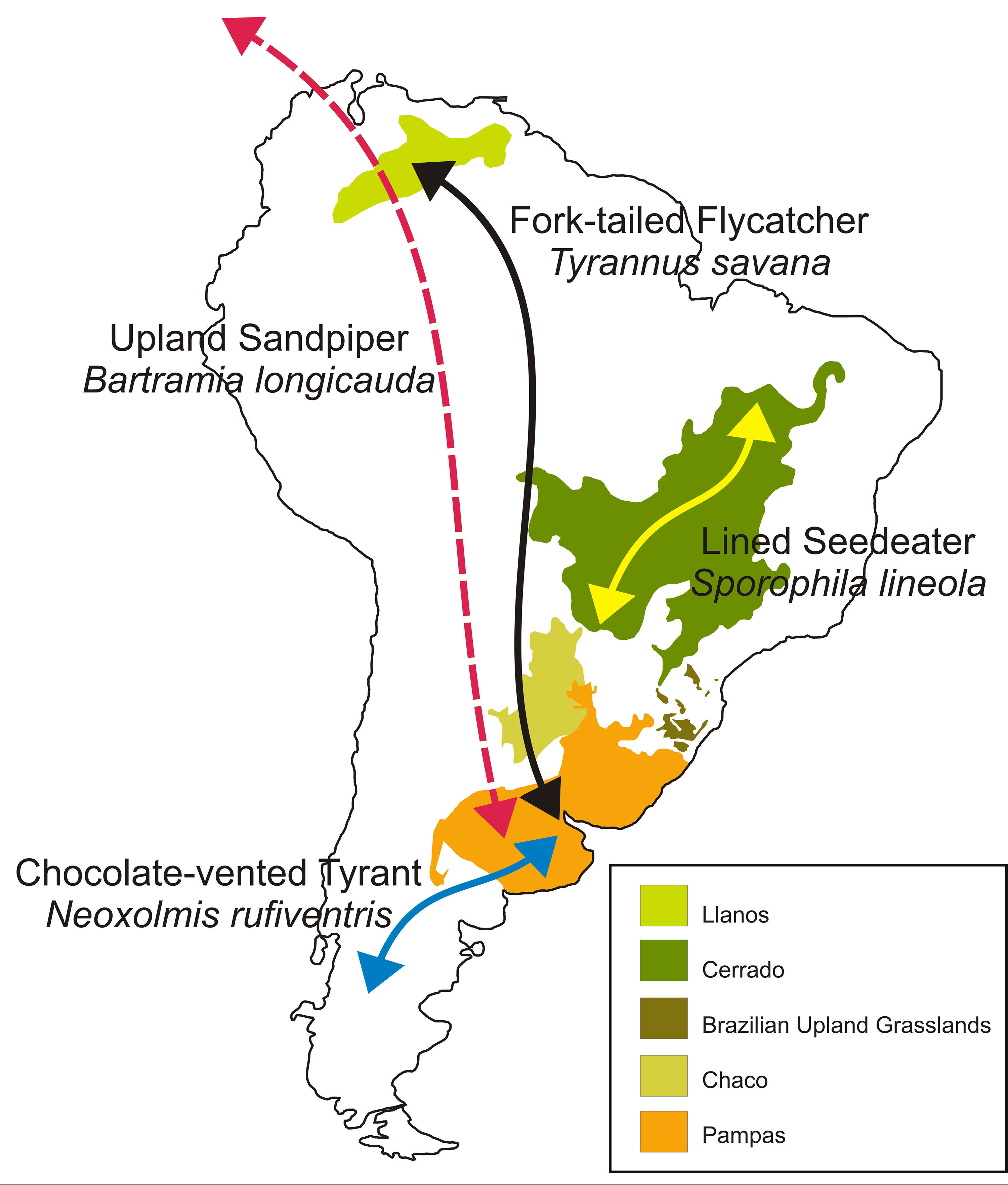
A brief introduction to South America's grassland migrantsFor the purposes of this discussion, we will focus on migratory birds occurring in three of South America's largest grasslands: (1) south-temperate grasslands of southeastern South America, including the upland grasslands (i.e., “campos de altitude”) of southern Brazil, the Pampas of the Uruguay, Paraná-Paraguay and La Plata River basins, and the wet grasslands of the Chaco of northern Argentina and southern Paraguay (Fig. 1; for a review of these grasslands and their birds, see Azpiroz et al., 2012), (2) the campo grasslands of the Cerrado ecoregion at the center of the continent (Fig. 1), and (3) the Llanos of the Orinoco River Basin in eastern Colombia and Venezuela (Fig. 1). Birds representative of several migratory systems occur in these grasslands:
- (1)
South American temperate-tropical migrants (sensuJoseph, 1997), which breed in south-temperate grasslands and overwinter in campo Cerrado grasslands (e.g., Chestnut Seedeater, Sporophila cinnamomea; Tawny-bellied Seedeater, Sporophila hypoxantha; Tropeiro Seedeater, Sporophila beltoni; Repenning and Fontana, 2016), or more rarely, in the Llanos (e.g., Blue-black Grassquit, Volatinia jacarina; Fork-tailed Flycatcher, Tyrannus savana, Jahn et al., 2013).
- (2)
South American cool-temperate migrants (sensuJoseph, 1997), which breed in Patagonian steppe and overwinter in south-temperate grasslands (e.g., Chocolate-vented Tyrant, Neoxolmis rufiventris; Gray-bellied Shrike-Tyrant, Agriornis micropterus).
- (3)
Intra-tropical migrants, which breed, migrate and overwinter in tropical grasslands, including campo Cerrado grasslands (e.g., Lesser Elaenia, Elaenia chiriquensis; Guaraldo et al., 2016; Lined Seedeater, Sporophila lineola; Silva, 1995).
- (4)
Nearctic-Neotropical migrants, which breed in the grasslands or arctic tundra of North America and overwinter in the Pampas (e.g., Upland Sandpiper, Bartramia longicauda; Bobolink, Dolichonyx oryzivorus; Renfrew et al., 2013).
, between South American grasslands and Patagonian steppe (i.e., Chocolate-vented Tyrant), and between South American grasslands and North America (i.e., Upland Sandpiper).")
Examples of bird migration between major grasslands of South America. Arrows represent general direction of migration of species that move between South American grasslands (i.e., Fork-tailed Flycatcher and Lined Seedeater), between South American grasslands and Patagonian steppe (i.e., Chocolate-vented Tyrant), and between South American grasslands and North America (i.e., Upland Sandpiper).
Admittedly, such a categorization does not fully describe the full diversity of avian migration in South America's grasslands, since some species fit into more than one category. For example, Chaco Pipit (Anthus chacoensis) migrates wholly within south-temperate grasslands (Casañas et al., 2007). Additionally, partial bird migration, in which only some populations of a species are migratory, is common in South America (Chesser, 1994). Yet, this scheme provides a framework within which to draw comparisons and discuss research and conservation priorities.
Of 288 grassland bird species in South America, 46 (16%) are migratory (including Nearctic-Neotropical migrants, Chesser, 1994; Vickery et al., 1999). To the best of our knowledge, the South American grasslands with the largest prevalence of migratory species are the south-temperate grasslands of southeastern South America. Of 109 species that occur in those grasslands, 33 (30%) are migratory (including Nearctic-Neotropical migrants; Azpiroz et al., 2012). Hereafter, we focus on birds that migrate wholly within South America.
Primary threats to South America's grasslands and their migrantsSouth America's grasslands face a broad suite of human impacts, resulting in greater pressure on natural resources such as water and less space for native species. Large-scale agriculture and overgrazing by cattle are arguably the greatest threats. Extensive planting of soy (Glycine max), rice (Oryza spp.), non-native grasses and trees, and widespread burning especially threaten the grasslands of the Cerrado and Atlantic Forest biomes (Silva and Bates, 2002; Klink and Machado, 2005; Silva et al., 2006; Repenning et al., 2010), which are among the most threatened Neotropical grasslands (Stotz et al., 1996; Azpiroz et al., 2012).
In the Pampas, extensive planting of soy, rice and wheat (Triticum spp.) and of exotic tree species such as Eucalyptus sp. (Krapovickas and Di Giacomo, 1998; Dias et al., 2013) have greatly reduced native habitat (reviewed by Azpiroz et al., 2012), leading to significant changes in the composition of grassland bird communities (Dias et al., 2013). Replacement of cattle-grazing cropping systems by continuous cropping systems (Medan et al., 2011) has led to overgrazing by cattle, fire and erosion (Berretta, 2001). In Argentina, Uruguay and southern Brazil, avian richness and density – especially that of grassland specialists – has diminished as grassland habitat has been converted to crop monocultures (Azpiroz and Blake, 2009; Codesido et al., 2013; Silva et al., 2015). Invasion by the non-native South African Lovegrass (Eragrostis plana; Carvalho and Batello, 2009) and other exotic species (Medan et al., 2011) are major threats. The use of agrochemicals, which pose a threat to multiple species, is widespread (Rivera-Milán et al., 2004; Zaccagnini, 2004), and several species are trapped for the pet trade (e.g., Sporophila seedeaters) many of which are classified as ‘Globally Threatened’ (BirdLife International, 2017). In Venezuela, threats to grassland migratory birds include use of toxic pesticides (e.g., Bobolink; Renfrew et al., 2013). Activities directly associated with crop monocultures, such as the persecution of migratory birds as agriculture pests (e.g., Bobolink; Basili, 1997) translates to additional pressures on their populations.
Currently, 12 grassland bird species migrating within South America are classified as ‘Globally Threatened’ or ‘Near Threatened’ (BirdLife International, 2017). Six species are considered Near-threatened (Hudson's Canastero, Asthenes hudsoni; Bay-capped Wren-spinetail, Spartonoica maluroides; Bearded Tachuri, Polystictus pectoralis; Dark-throated Seedeater, Sporophila ruficollis; Rufous-rumped Seedeater, Sporophila hypochroma; Black-bellied Seedeater, Sporophila melanogaster; BirdLife International, 2017). Another six species are considered globally threatened (Marsh Seedeater, Sporophila palustris; Chestnut Seedeater, S. cinnamomea; Tropeiro Seedeater, S. beltoni; Ibera Seedeater, Sporophila iberaensis; Strange-tailed Tyrant, Alectrurus risora; Pampas Meadowlark, Leistes defilippii; BirdLife International, 2017). The last two species have had their migratory behavior disrupted due to habitat loss and population decline. The Strange-tailed Tyrant has historically bred from southern Paraguay to central Argentina (Collar et al., 1992), overwintering from northern Argentina (Di Giacomo and Di Giacomo, 2004) to central Brazil (Pacheco and Gonzaga, 1994). At present, only small populations are resident in southern Paraguay and northern Argentina (Formosa, Chaco and Corrientes provinces). The Pampas Meadowlark was historically a partial migrant, with most of the breeding population located in southern Buenos Aires Province, Argentina, and which migrated north to southern Brazil and Uruguay (Tubaro and Gabelli, 1999). At present, a large fraction of the population remains resident throughout the year in southern Buenos Aires Province (Gabelli et al., 2004), isolated from another small resident population in western Uruguay (Azpiroz, 2005).
Although numerous recommendations have been made for grassland bird conservation in South America (reviewed by Azpiroz et al., 2012), our lack of knowledge on the basic natural history of grassland migratory birds on the continent has hindered meaningful, population-level or even species-specific planning for these species. We still lack such basic information about where specific populations overwinter, where their migratory routes are located, and even which species migrate. In summary, the lack of basic knowledge on grassland migratory bird biology, combined with novel and constantly changing impacts on the grasslands they inhabit requires an urgent, inter-disciplinary, and geographically broad research and conservation effort.
Carry-over effects, connectivity and migratory networksEffective conservation planning for migratory organisms first requires understanding their behavioral and physiological ecology throughout the entire annual cycle, since these have the potential to ultimately affect a population's growth rate (Harrison et al., 2011; Marra et al., 2015). For example, the events that occur within the life of an individual bird may be driven by mechanisms operating within a given season (e.g., nest predation driven by microhabitat near the nest), as well as between seasons (e.g., winter habitat quality affecting body condition and reproductive success in summer; e.g., Marra et al., 1998; Studds and Marra, 2005; Norris and Taylor, 2006). Studying such ‘carry-over effects’ can be challenging, since a migratory organism's annual cycle may involve events occurring across hundreds to thousands of kilometers. However, carry-over effects are important to understand, since they have the potential to negatively impact migratory bird productivity (e.g., Inger et al., 2010).
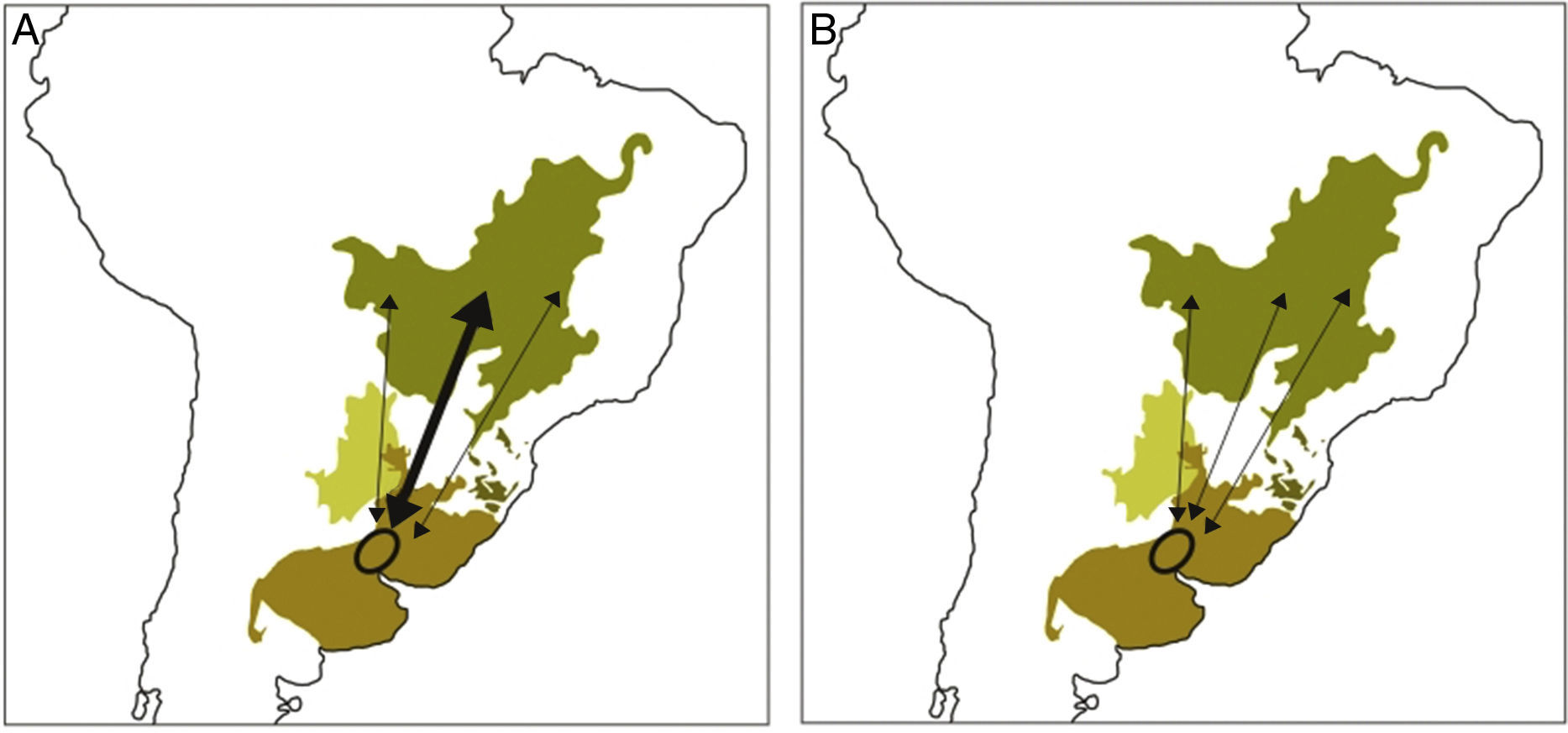
Understanding how processes such as carry-over effects scale up from the individual to the population level (e.g., consequences of individual winter habitat quality on demographic rates during the breeding season) requires information on migratory connectivity (i.e., the extent to which individuals from the same breeding area migrate to the same nonbreeding area and vice versa; Webster et al., 2002; Norris and Marra, 2007). For example, a hypothetical breeding population of Tawny-bellied seedeaters may exhibit strong migratory connectivity, such that most or all individuals of a breeding population overwinter in the central Cerrado grasslands (Fig. 2A). Conversely, that population may be weakly connected among seasons, with most individuals overwintering across most of the Cerrado ecoregion (Fig. 2B). If, as in the first case, most Tawny-bellied seedeaters from that breeding population overwinter in the central Cerrado and that region experiences serious habitat degradation, that breeding population may be negatively affected (i.e., if carry-over effects from winter to breeding occur). In North America, populations of Cerulean Warblers (Setophaga cerulea) with the strongest migratory connectivity between breeding and wintering sites are those that have experienced the steepest declines in their populations (Jones et al., 2008).
 showing strong migratory connectivity (A), in which most individuals from the breeding population move to the same wintering location in the central Cerrado ecoregion (wide arrow), with smaller proportions of the breeding population moving to different wintering locations elsewhere in the Cerrado (narrow arrows), and weak migratory connectivity (B), in which similar proportions of individuals from the breeding population migrate to several wintering locations.")
Hypothetical breeding population of Tawny-bellied Seedeater (circle) showing strong migratory connectivity (A), in which most individuals from the breeding population move to the same wintering location in the central Cerrado ecoregion (wide arrow), with smaller proportions of the breeding population moving to different wintering locations elsewhere in the Cerrado (narrow arrows), and weak migratory connectivity (B), in which similar proportions of individuals from the breeding population migrate to several wintering locations.
Potentially more challenging is research on processes operating within a migratory network, composed of breeding, stopover and overwintering sites used by multiple populations (Taylor and Norris, 2010; Betini et al., 2015). For example, habitat loss at a given wintering site can affect the connectivity of two or more breeding populations. In the case of our hypothetical population of Tawny-bellied seedeaters with strong migratory connectivity (Fig. 2A), major habitat loss in the central Cerrado, where a large proportion of a breeding population overwinters, could potentially lead to weaker connectivity of that population to that wintering site and drive a larger proportion of them to overwinter in other regions of the Cerrado (if habitat is available and they have the intrinsic flexibility to do so; Taylor and Norris, 2010).
Given the challenges of collecting adequate information on carry-over effects at the individual level, and on connectivity and migratory networks at the population level, it is not surprising that full annual cycle research of migratory bird populations is rare, even in the relatively well-studied Nearctic-Neotropical and Palearctic-Paleotropical migratory systems. Logistic concerns aside, many studies do not include analysis of movement, due in part to a mismatch in the temporal or taxonomic scale at which movement occurs and the scale at which most biodiversity research is focused (Nathan et al., 2008; Jeltsch et al., 2013). Indeed, much research is focused on the breeding season (Marra et al., 2015) and at the species and community levels, whereas movement ecology is primarily concerned with processes occurring at the individual level and beyond a single season (Jeltsch et al., 2013).
Movement ecology research to conserve South American grassland migrantsHow can movement ecology research best contribute to conservation of South America's grassland migratory birds? We suggest that addressing the following questions is fundamental for understanding how to best conserve a given grassland migratory bird population. Some of these questions may not be immediately feasibly addressed, due to lack of funding, training, or logistical hurdles. Yet, considering these questions, in combination with a well-defined rationale about which questions are worth the time and money needed to answer them (McGowan et al., 2016), can aid in deciding how to prioritize current and future research objectives and conservation planning.
Which populations of a given species migrate? Although migratory patterns at the species level are increasingly better described, due to basic biogeographic research and growing, publicly available online databases (e.g., GBIF, eBird, WikiAves), we still have only a rudimentary understanding of migration of South America's grassland birds at the population level (Azpiroz et al., 2012), as is generally true of most bird species that migrate within South America (Jahn et al., 2009). Because partial migration is so common on the continent (Stotz et al., 1996), detecting migration in a given population is not straightforward (e.g., Sagario et al., 2014). Nevertheless, standardized, systematic and long-term monitoring, including international collaborations, promise unprecedented discoveries about how and why birds migrate on the continent (Jahn et al., 2009).
What is the migratory connectivity of grassland migrants? Because a basic grasp of population migratory connectivity is imperative for understanding how individual attributes drive population-level dynamics, evaluating the level of migratory connectivity (or lack of knowledge thereof) is key during the conservation planning process (Martin et al., 2007; Finch et al., 2017). Furthermore, a basic grasp of connectivity across several populations offers an opportunity to develop conservation plans that take into consideration mechanisms operating throughout a species’ migratory network (i.e., across multiple populations; Taylor and Norris, 2010).
Although studying migratory connectivity can be challenging, for example due to lack of isotope maps for many regions, new techniques are constantly being developed for modeling connectivity, including those for species which occur in areas with few isotopic measurements (Pekarsky et al., 2015). However, a detailed description of the molt cycle is still lacking for many migratory bird species in South America, which precludes using feather isotope analysis to study connectivity. Other tissues, such as claws (e.g., Guaraldo et al., 2016) can also be sampled to study the connectivity of South America's migratory birds. It is therefore imperative for researchers to collect tissue samples, such as claws and feathers, as well as detailed molt data from migratory birds in South America. Additionally, as tracking devices become smaller and lighter, their use is a viable option to understand the connectivity of many migratory species (Kays et al., 2015).
What are the individual attributes of grassland migrants throughout the year? Identifying appropriate management actions and the scale at which to implement them requires an understanding of individual attributes, such as timing of spring or fall migration (Allen and Singh, 2016; McGowan and Possingham, 2016) and dispersal (Jønsson et al., 2016), and potential for molt-migration (Guaraldo et al., 2016). One might ask: How, to where and why does a grassland migratory bird move (Nathan et al., 2008; Jeltsch et al., 2013)? For example, collecting data on timing of migration (e.g., dates of arrival on wintering and breeding grounds) of Sporophila seedeaters, combined with data on timing of seeding phenology (since their migration is adapted, potentially, to track seed availability; Remsen and Hunn, 1979; Silva, 1999), can provide vital information on identifying when and where bottlenecks to their survival and condition exist.
Though logistically challenging, collecting individual movement data can be accomplished by employing recently developed technologies and techniques, such as miniature tracking devices (reviewed by Kays et al., 2015), which offer an unsurpassed ability to understand the annual cycle of even relatively small migrants. Juvenile dispersal in particular has been difficult to study, yet emerging techniques promise an opportunity to begin quantifying dispersal, even for small organisms. Given its key role in colonization and the mixing of gene pools within and between populations, dispersal research is an exciting and rapidly growing field that promises important breakthroughs (Jønsson et al., 2016). In summary, linking movement data with parallel data on physiology (e.g., energetic condition; Peig and Green, 2009), molt timing (Barta et al., 2008), demographics (age and sex), and nest and/or adult survival (e.g., with mark-recapture models, Hostetler et al., 2015) is vital for advancing grassland migratory bird conservation.
What carry-over effects exist in the annual cycle of grassland migrants? A basic understanding of if and how carry-over effects operate in a given population is vital to streamlining conservation action, especially if a population exhibits strong migratory connectivity, since interactions between seasons can be magnified in such a population (Calvert et al., 2009). As in our previous example of a hypothetical breeding population of Tawny-bellied seedeaters exhibiting strong migratory connectivity (Fig. 2A), habitat loss where they overwinter potentially translates to negative impacts during the breeding season via carry-over effects. Although carry-over effects are challenging to study, novel analytical techniques, such as state-dependent modeling, can help distinguish changes in population size as a result of carry-over effects (e.g., reduced reproductive output due to poor winter habitat quality) versus other sources (e.g., mortality; Harrison et al., 2011).
What role do grassland migrants play in their ecosystems? Studying how the species of interest impacts its ecosystem is a key step in identifying conservation priorities, since using animal movement data for conservation planning permits pinpointing the ecosystem services provided by movement and therefore management at the landscape level (Jeltsch et al., 2013; Mitchell et al., 2013; Allen and Singh, 2016). Additionally, given that at least some of South America's migratory grassland birds occupy more than one grassland type (Fig. 1), they may not only play an important role within one, but among several grassland types. As a result, losing such migratory populations may not only alter ecosystem services locally, but regionally. Thus, international research collaboration on grassland bird migration may not only serve to understand migrants, but their role within their ecosystem(s) throughout the year, and how they promote ecosystem function at different spatiotemporal scales.
ConclusionsIn summary, incorporating a movement ecology perspective to basic and applied research on South American grassland migratory birds promises a more comprehensive understanding of their biology across the year, and therefore vital information for conserving and managing their populations. Combining molecular, ecological, behavioral and physiological approaches with movement ecology's theoretical perspective and analytical tools offers a much broader appreciation of the lives of grassland birds. Given the diverse set of threats to South America's grasslands and their migrants, from fragmentation of habitat locally to climate change at a larger scale, such interdisciplinary research is badly needed and necessary to address such pressing questions as: What are the primary drivers of population demographic rates of grassland migratory birds? And, given that migratory birds on other continents have been negatively impacted by climate change (e.g., Both et al., 2006; Wilcove and Wikelski, 2008; Jones and Cresswell, 2010), which populations of grassland migratory birds in South America are most vulnerable to climate change?
Accomplishing this will require moving away from breeding-centric research toward that on the full annual cycle of individuals. Its success will no doubt depend heavily on strong research collaborations across national and international political boundaries, as is true of many conservation initiatives (e.g., ‘Alianza del Pastizal’ initiative, http://www.alianzadelpastizal.org).
Only through such a spatially and temporally broader research focus and through the creation of strong international research partnerships will we achieve a basic understanding of the complex yet fascinating lives of grassland migratory birds throughout their annual cycle, including the influence of such processes as carry-over effects on the survival of individuals and ultimately, species. Such international research initiatives are a prerequisite to the creation of management and conservation actions at an appropriate spatial scale, since many species (e.g., Fork-tailed Flycatcher; Jahn et al., 2013) spend different parts of the year in different countries. The time is ripe for researchers to incorporate movement ecology theory and tools to further our understanding of grassland migratory bird biology in South America, and consequently our ability to conserve these unique animals.
We dedicate this paper to the late Peter Vickery. This manuscript benefitted greatly from the comments of two anonymous reviewers. AEJ thanks the Fundação de Amparo à Pesquisa do Estado de São Paulo (2012/17225-2, 2013/19116-9), ASDG thanks Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT, PICT 2014-3397) and CSF thanks Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq 303318/2013-9, 457475/2012-2) for support.

